Download

1 / 33
340 likes | 408 Views
Enzymes are essential biological catalysts that accelerate reaction rates by lowering activation energies. Learn about their structure, properties, and mechanisms of activity in living systems.

E N D
Enzymes as Catalysts • Living processes consist almost entirely of biochemical reactions. In the absence of catalysts these reactions would not occur fast enough to sustain life. • To proceed at a viable rate, most chemical reactions require an initial input of energy. Heat is used for providing the energy in the non-living processes. Heat increases the likelihood of the collision of the vibrating molecules. A chemical reaction occurs when the colliding molecules possess a minimum amount of energy, i.e. activation energy. Increasing the concentration of the reactants also increases the likelihood of their collision and the formation of product. • Living processes, however, suffer from two limitations: 1- elevated temperatures may be harmful to the delicate biological structures and 2- reactant concentrations are often quite low. • One of the most important functions of proteins is their role as catalysts. • Living systems benefit from enzymes as catalysts. Like other catalysts, enzymes enhance reaction rates by lowering activation energies. They have no effect on the position of reaction equilibria.
Enzyme Properties • Enzymes are known to have 3 critical properties: the rate of enzymatically-catalyzed reactions is remarkably high, the enzymes are highly specific to the reactions they catalyze, and enzymes can be regulated (this is crucial, since living organisms must conserve energy and raw materials). • An enzyme is a globular protein which functions as a biological catalyst, speeding up reaction rate by lowering activation energy without being affected by the reaction it catalyses. • Enzymes (from the Greek enzymos, “leavened”) are the chemical catalysts of biological systems. • Enzymes function in aqueous solutions under very mild conditions of temperature and pH, unlike many catalysts used in organic chemistry. • Some enzymes require no chemical groups for activity other than their amino acid residues. Others require an additional chemical component called a cofactor. Cofactors can be inorganic ions, or complex organic or metallo-organic molecules called coenzymes.
Cofactors and Coenzymes Coenzymes usually act as transient carriers of specific functional groups. Most are derived from vitamins. Some enzymes require both a coenzyme and one or more metal ions for activity. A coenzyme or metal ion that is very tightly or even covalently bound to an enzyme protein is called a prosthetic group. A complete, catalytically active enzyme together with its bound coenzyme and/or metal ion is called a holoenzyme. The protein part of such an enzyme is called the apoenzyme or apoprotein. Many enzymes are modified by phosphorylation or other processes. Modifications often are used to regulate enzyme activity.
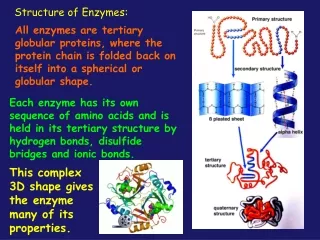
Structure of enzymes Enzymes Complex or holoenzymes (protein part and nonprotein part – cofactor) Simple (only protein) Cofactor Apoenzyme (protein part) Coenzyme -large organic molecule -loosely bound to apoenzyme Prosthetic groups • usually small inorganic molecule or atom; • usually tightly bound to apoenzyme Example of metalloenzyme (prosthetic group): carbonic anhydrase contains zinc
Enzymes as catalysts of biological reactions may accelerate reactions by a millions fold:
Coenzymes act as group-transfer reagents: Hydrogen, electrons, or groups of atoms can be transferred. Coenzyme classification: Metabolite coenzymes - synthesized from common metabolites Vitamin-derived coenzymes - derivatives of vitamins ATP Examples of metabolite coenzymes: ATP can donate phosphoryl group S-adenosylmethionine donates methyl groups in many biosynthesis reactions S-adenosylmethionine
Vitamin-Derived Coenzymes: NAD+ and NADP+ • Most vitamins must be enzymatically transformed to the coenzyme. Vitamins are required for coenzyme synthesis (in mammals) and must be obtained from nutrients. Deficit of vitamin and as result correspondent coenzyme results in the disease. • Nicotinic acid (niacin) and nicotinamide are precursor of NAD and NADP. • NAD and NADP are coenzymes for dehydrogenases.
Vitamin-Derived Coenzymes: FAD and FMN • Flavin adenine dinucleotide (FAD) and Flavin mononucleotide(FMN) are derived from riboflavin (Vitamin B2). • Flavin coenzymes are involved in oxidation-reduction reactions. FMN (black), FAD (black/blue)
Vitamin-Derived Coenzymes: Thiamine Pyrophosphate (TPP) • TPP is a derivative of thiamine (Vit B1). • TPP participates in reactions of: (1) Oxidative decarboxylation(2) Transketo-lase enzyme reactions.
Vitamin-Derived Coenzymes: Pyridoxal Phosphate (PLP) • PLP is derived from Vit B6 family of vitamins. PLP is a coenzyme for enzymes catalyzing reactions involving amino acid metabolism (isomerizations, decarboxylations, transamination).
Enzyme Active Site Under biologically relevant conditions, uncatalyzed reactions tend to be slow because most biological molecules are quite stable in the neutral pH, mild temperature, aqueous environment inside cells. Enzymes greatly increase the rates of biological reactions by providing a specific environment within which a reaction can occur more rapidly. Enzyme-catalyzed reactions take place within the confines of a pocket on the enzyme called the active site. The reactant molecule is referred to as the substrate. The surface of the active site is lined with amino acid residues with substituent groups that bind to the substrate and catalyze its chemical transformation. Often, the active site encloses the substrate, sequestering it from solution. Substrate usually is a relatively small molecule. Enzyme is large protein molecule. Therefore substrate binds to specific area on the enzyme. Substrate interacts (weak linkages of non-covalent bonds) with only 3-5 amino acid residuesthat could be far apart in sequence.
Free Energy and Reaction Rate Any reaction, such as S P, can be described by a reaction coordinate diagram, in which the free energy change during the reaction is plotted as a function of the progress of the reaction. The free energy change (∆G’0) (and equilibrium position) of the reaction is determined by the difference in ground state free energies of S and P.The rate of the reaction is dependent on the height of the free energy barrier between S and P. At the top of this hump is the transition state. The transition state is not a chemical species with any significant stability, and should not be confused with a reaction intermediate. Rather it is a fleeting molecular moment in which events such as bond breakage, bond formation, and charge development have proceeded to the point at which decay to either substrate or product is equally likely. The difference between the energy levels of the ground state and the transition state is the activation energy, ∆G‡. The rate of the reaction is inversely and exponentially proportional to the value of ∆G‡.
Activation Energy (∆G‡) and Reaction Rate The rate of a chemical reaction is determined by the concentration of the reactant(s) and by a rate constant usually denoted by k. For the unimolecular reaction S P, the rate (or velocity) of the reaction, V (i.e. the amount of S that reacts per unit time) is expressed by a rate equation, V = k[S]. In this reaction, the rate depends only on the concentration of S. This is a first-order reaction. The factor k is a proportionality constant that reflects the probability of a reaction under a given set of conditions (pH, temperature, etc.). Here, k is a first-order rate constant and has the units of reciprocal time (s-1). If a reaction rate depends on the concentration of two different compounds, or if the reaction is between two molecules of the same compound, then the reaction is second-order and k is a second-order rate constant, with units of M-1s-1. The rate equation then becomes V = k[S1][S2]. From physical chemistry, it can be derived that the magnitude of a rate constant is inversely and exponentially related to the activation energy, ∆G‡. Thus, a lower activation energy means a faster reaction rate. E + S ⇄ ES ⇄ EP ⇄ E + P Binding energy provided by the interaction of the enzyme with the S at transition state contributes strongly to lowering the activation energy of the reaction, and accelerating its rate.
Specificity of Enzymes • Enzymes not only bring about enhancements in reaction rates they are also very specific, readily discriminating between substrates with quite similar structures. The rate enhancements observed for enzymes come from two distinct but interwoven parts. First, catalytic functional groups on an enzyme react with a substrate and lower the activation energy barrier for the reactions by providing an alternative, lower-energy reaction path. • Second, noncovalent binding interactions between the substrate and enzyme release a small amount of free energy with each interaction that helps lower the energy of the transition state. The energy derived from enzyme-substrate interaction is called the binding energy, ∆GB. • Structural complementarity is responsible for the high specificity of enzyme reactions. The idea that the enzyme and substrate are complementary to one another (like a lock and key) was first proposed by the organic chemist, Emil Fisher, in 1894. • However, precise complementarity between an enzyme and its substrate would be counterproductive to efficient catalysis. The enzyme must be more complementary to the reaction transition state than to the substrate per se for efficient catalysis to occur.
Binding Energy and Reaction Rate • Some weak interactions are formed in the ES complex, but the full complement of such interactions between the S and E is formed only when the substrate reaches the transition state. The free energy (binding energy) released by the formation of these interactions partially offsets the energy required to reach the top of the energy hill. The summation of the unfavorable (positive) activation energy ∆G‡ and the favorable (negative) binding energy ∆GB results in a lower net activation energy. Even on the E, the transition state is not a stable species but is a brief point in time that the S spends atop an energy hill. The enzyme-catalyzed reaction is much faster than the uncatalyzed process because the hill is much smaller. The important point is that weak binding interactions between the E and the S provide a substantial driving force for enzymatic catalysis. • Binding energy enhances the reaction rate via 1- entropy reduction (the restriction in the motions of substrates and 2- desolvation (formation of weak bonds between the E and S results in the desolvation of the substrate, i.e. removal of bound water molecules) and 3- catalytic group alignment. Binding energy also gives an E its specificity (i.e. ability of E to discriminate between its S and a competing molecule with a similar structure.
Specificity of Enzymes • Enzymes have varying degrees of specificityfor substrates. • Enzymes may recognize and catalyze: A single substrate (Absolute specificity) A group of similar substrates which have the same bond type (Group specificity, i.e. relative specificity) A particular type of bond (Linkage specificity) The transformation only (i.e. Stereospecificity: substrates which are in certain geometrical configuration, e.g. cis- or trans-)
Lock-and-Key Model of Enzyme Action In the lock-and-key model of enzyme action: the active site has a rigid shape, this explains enzyme specificity, it explains the loss of activity when enzymes denature.
Induced-Fit Model of Enzyme Action Catalytic group alignment: Enzymes typically undergo changes in conformation when the S binds (induced by multiple weak interactions with the S). The alignment of catalytic functional groups is referred to as induced fit, and it serves to bring specific functional groups on the enzyme into the proper position to catalyze the reaction. • In the induced-fit modelof enzyme action: - the active site is flexible, not rigid - the shapes of the enzyme, active site, and substrate adjust to maximize the fit, which improves catalysis. - there is a greater range of substrate specificity. • This model is more consistent with a wider range of enzymes.
Active site contains functional groups (-OH, -NH, -COO etc) Binds substrates through multiple weak interactions (noncovalent bonds)
Principles of Enzyme Classification Common names are formed by adding the suffix –ase to the name of substrate (e.g. cellulase catalyzes the hydrolysis of cellulose). Common names don’t describe the chemistry of the reaction. Trivial names don’t give information about the substrate, product or chemistry of the reaction (e.g. catalase). Biochemists by international agreement have adopted a system for naming and classifying enzymes based on the type of reaction catalyzed. Each enzyme is assigned a four-part classification number and a systematic name, which identifies the reaction it catalyzes.. Hexokinase catalyzes: Glucose + ATPglucose-6-P + ADP
Enzyme Commission Classification All enzymes are classified into six categoriesaccording to the type of reaction they catalyze. For example the enzyme known commonly as hexokinase is formally ATP:glucose phosphotransferase. Its Enzyme Commission number is EC 2.7.1.1, in which the first number (2) denotes the class name (transferase); the second number (7), denotes the subclass (phosphotransferase); the third number (1), indicates the sub-sub class , a phosphotransferase with a hydroxyl group (alcohol) as acceptor; and the fourth number (1), is specific name: ATP,D-HEXOSE-6-PHOSPHOTRANSFERASE (Hexokinase).
The First Class of Enzymes EC 1.Oxidoreductases Include:oxidases, peroxidases and dehydrogenases Catalyze: oxidation-reduction reactions Examples:Lactate dehydrogenase, Glucose oxidase, Peroxidase, Catalase, Phenylalanine hydroxylase.
EC 2.TransferasesIncludes: esterases, peptidases and glycosidasesCatalyze:group transfer reactions.Transfer a functional group (e.g. methyl or phosphate) between donor and acceptor molecules.Examples: Transaminases, Phosphotransferases (Kinases), Transmethylases, Transpeptidases, Transacylases The Second Class of Enzymes
EC 3. HydrolasesCatalyze:hydrolysis reactions where water is the acceptor of the transferred group(Add water across a bond).Example: Protein hydrolyzing enzymes (Peptidases), Carbohydrases (Amylase, Maltase, Lactase), Lipid hydrolyzing enzymes (Lipase), Deaminases, Phosphatases. The Third Class of Enzymes
EC 4. LyasesCatalyze:lysis of a substrate, cleave various bonds, generating a double bond in a nonhydrolytic, nonoxidative elimination.Add water, ammonia or carbon dioxide across double bonds, or remove these elements to produce double bonds.Example: Fumarase, Carbonic anhydrase The Fourth Class of Enzymes
EC 5. Isomerases Catalyze:isomerization reactions (e.g. L to D and Cis to Trans isomerizations, Mutase reactions such as shifts of chemical groups).Example: Isomerase and Mutase The Fifth Class of Enzymes
EC 6.Ligases(synthetases) The Sixth Class of Enzymes • Catalyze:ligation, or joining of two substrates. • Require chemical energy (e.g. ATP) • Example:Acetyl~CoA carboxylase,Glutamine synthetase