Download

1 / 24
240 likes | 272 Views
Chapter 3. Membrane Transport. 3.1 Overview of membrane transport. 1) Membranes facilitate compartmentalization. Lipophilicity and membrane permeability. Ps : permeablity coefficient Mr : melecular mass. 2) Selective permeability of biological membranes derives from

E N D
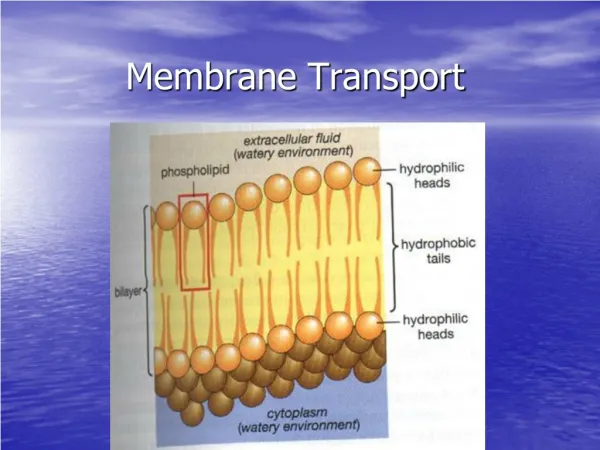
3.1 Overview of membrane transport 1) Membranes facilitate compartmentalization
Lipophilicity and membrane permeability Ps : permeablity coefficient Mr : melecular mass
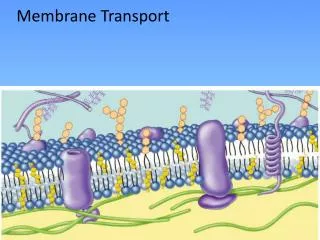
2) Selective permeability of biological membranes derives from transport systems that contain integral membrane proteins KAT1 KAT1 D95-AP, KAT1K128-AP fusion proteins
3) Membrane transport underlies many essential cell biological processes. ▶ Turgor generation: Ions (K+ in cytosol, K+, Na+ and Cl-, malate in vacuoles) ▶ Nutrient acquisition: NH4+, NO3-, H2PO4-, SO4- , B, Zn, Cu, Fe, ▶ Waste product excretion: H+, oxalate, ▶ Metabolite distribution: Sucrose and amino acids to pheloem. ▶ Compartmentalization of metabolites: transport for conversion ex) amyloplast(glucose6-p), matrix of mitochondria (ADP/ATP, NADPH/NADP+ ratios are high)
▶ Energy transduction: H+ gradient and ATP synthesis ▶ Signal transduction: cytosolic free Ca2+
3.2 Organization of transport at plant membranes • Metabolically coupled H+-pumps underlie • a proton-based energy economy in plants
Box3-3. The electrochemical potential of a solute is defined by differences in concentration and charge.
Box3-4. What is a membrane potential? Normally, Vm= -150 mV across PM Vm= -20 mV for tonoplast Positive value indicates that energy input is required for uptake of solute ion.
2) Proton recirculation drives solute absorption and excretion by way of specific symporters and antiporters
3) Channels catalyze the movement of specific ions in the net direction of their electrochemical potential driving forces. K+, Ca2+, Cl- etc.
4) Turnover rates among classes of transport system vary, affecting protein abundance in membranes.
PM H+-ATPase Fig.3.2. SDS-PAGE of fractions obtained during purification of PM H+-ATPase from spinach leaves.
3.3.1. F-type H+-ATPases at the inner mitochondrial and thylakoid membrane are pumps that operate in reverse mode to synthesize ATP
Model for the synthesis of ATP by an F-type H+-pumping ATPase O: open, L: binds loosely, T: binds tightly
3.3.2. PM H+-ATPase performs a variety of physiological functions and is a P-type ATPase • A single peptide of about 100 kDa • It drive fluxes through other transporter systems(by proton motive force) • The activation is one of the earliest responses to auxin. • It is a critical in removing excess H+ from the cytosol.
3.3.3. PM H+-ATPase is encoded by a multigene family that exhibits tissue specific expression (at least 10 genes) Gene expression of AHA3
3.3.4 Plasma membrane H+-ATPase is regulated by an array of mechanisms. H+-ATPase Binding sites for H+ Phosphorylation of Aspartyl residue C-terminus cytosolic region Catalyzing dephosphorylation Autoinhibitory domain Fold back into the protein to accomplish inhibition
Fusicoccin A Toxin produced by fungus (Fusicoccum amygdali), a phathogen affecting peach and almond tree. Elicits an increase in guard cell turgor, thereby opening stomata and causing leaves to wilt. Stimulates the P-type H+-ATPase
Autoinhibitory domain in C-terminal Phosphorylation of a serine 14-3-3 proteins A family of soluble signal transduction proteins Bind to phosphorylated serinyl residue Relieving C-terminal autoinhibition Binding of two 14-3-3 proteins
3.3.5 H+:ATP stoichiometry determines poise. nH+i + ATP nH+o +ADP + Pi F-type H+-ATPase: n=3 P-type H+-ATPase: n=1