Download

1 / 71
840 likes | 2.37k Views
Recombinant DNA cloning technology. Chapter 7. DNA cloning. To obtain large amounts of pure DNA Procedure Isolate DNA Use restriction enzymes to cut DNA Ligate fragments into a cloning vector Transform recombinant DNA into a host to replicate the DNA and pass copies into progeny.

E N D
Recombinant DNA cloning technology Chapter 7
DNA cloning • To obtain large amounts of pure DNA • Procedure • Isolate DNA • Use restriction enzymes to cut DNA • Ligate fragments into a cloning vector • Transform recombinant DNA into a host to replicate the DNA and pass copies into progeny.
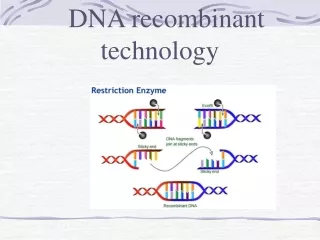
Restriction Enzymes • Recognize a specific DNA sequence (restriction site) • Break a phosphodiester linkage between a 3’ carbon and phosphate • Used for • Create DNA fragments for cloning • Analyze positions of restriction sites in cloned or genomic DNA
Restriction Enzymes • Are found naturally in bacteria as a defense against vital DNA. • Restriction sites are methylated in bacteria, and thus protected.
Restriction Enzymes • Are denoted by three letter names derived from the bacterial strain they originate from.
Restriction Enzymes • They usually are palindromes of 4-, 6- or 8-base pairs. • Based on the probability, a specific short DNA sequence occur more frequently than a long one. • In 50% GC content, each base has a ¼ chance of occurring at a position. • The frequency of a particular restriction site is (1/4)n.
Restriction Enzymes: EcoRI • EcoRI (“Echo R one”) is a commonly used enzyme. It was the first (one) restriction enzyme isolated from the “R” strain of E. coli. It demonstrates the usual type recognition site, a palindrome (the same on both strands, reading in opposite directions) EcoRI leaves a four base, 5’ overhang, sticky end.
Restriction Enzyme Sites • Sma I (from Serratia marcescens) cuts a palindrome to give blunt ends. • BamHI (from Bacillus amyloliquefaciens H) cuts to give a 5’ overhang. • PstI (from Providencia stuartii) cuts to give a 3’ overhang.
Restriction Enzymes • Blunt ends: both strands are cut at the same position. • Sticky ends: overhanging regions (3’ or 5’) are useful in cloning. They are complementary, thus anneal, DNA ligase can covalently link them.
Question 7.2 • A new RE from a bacterium cuts DNA into fragments that average 4096 bp long. This RE has a two-fold rotational symmetry. How many based pairs of DNA constitute the recognition site for the new enzyme? • Two-fold rotational symmetry: axis of symmetry through the midpoint. The base sequence from 5’ to 3’ on one DNA strand is the same as the base sequence from 5’ to 3’ on the complementary strand.
Answer 7.2 • The average length of fragments produced indicates how often the RE site appears, in this case every 4096 bp, there is an RE in the genome. • If DNA is composed of equal amounts of A, T, C, and G, then the chance of finding one specific basepair (AT, TA, GC, or CG) at a particular site is ¼. • Finding n number of specific basepairs is (1/4)n • 1/4096=(1/4)6
Question 7.3 • An endonuclease called AvrII cuts DNA whenever it finds the sequence 5’-CCTAGG-3’ 3’-GGATCC-5’ About how many cuts would AvrII make in human genome, which is about 3x109 bp long and about 40%GC.
Answer 7.3 5’-CCTAGG-3’ 3’-GGATCC-5’ The enzyme recognizes a sequence that has two GC bp, two CG bp, one AT bp, and one TA bp in a particular order. Since genome has 40% GC, the chance of finding a GC or a CG pair is 0.2 while the chance of finding an AT or a TA pair is 0.3. The chance of finding the 6 base pairs in this sequence is: P(AvrII site) = 0.2x0.2x0.3x0.3x0.2x0.2=0.000144 Number of sites in the genome = 0.000144 x (3 x 109) = 432,000.
Cloning DNA with the Restriction Enzyme EcoRI • A typical DNA cloning experiment requires that the DNA to be cloned (often called the “insert DNA”) and the vector (often a plasmid) both be cut with the same enzyme (or with two enzymes which produce compatible ends). The insert DNA and the vector are then mixed, and DNA ligase is used to join the molecules.
Cloning Vectors • Plasmid; • Phage; • Cosmid; • Shuttle; • Yeast Artifical Chromosomes (YACs) • Bacterial Artificial Chromosomes (BACs)
Plasmid Cloning Vectors • Derived from natural plasmids. • Plasmids are circles of dsDNA that include origin sequences (ori) needed for replication in bacterial cells. • E.coli plasmid vectors contains • An ori sequence required for replication in E.coli • A selectable marker, e.g., antibiotic resistance, to allow selection of cells harboring the plasmid. • At least one unique restriction enzyme cleavage site, so that DNA sequences cut with the RE can be spliced into the plasmid.
pUC19 (2,686-bp) • High copy number in E.coli, ~100 copies per cell (makes it easy to purify the plasmid); • Has a selectable marker ampR (indicates the success of transformation). • A cluster of unique restriction sites, called polylinker (multiple cloning site makes it easy to use different RE sites for cloning). • Polylinker is part of lacZ gene (B-galactosidase). If pUC19 plasmid is put in a lacZ- E. coli, cell will become lacZ+. • If DNA is cloned into the polylinker, lacZ is disrupted, no complementation occur.
Plasmid pUC 19 • The commonly used plasmid pUC19 (“puck 19”) is a small plasmid with the essential elements for a vector: An origin of DNA replication A dominant selectable marked (resistance to an antibiotic, ampicillin) And a cloning site, usually a polylinker with recognition sites for numerous restriction enzymes.
How to detect if DNA is cloned into the polylinker? • X-gal, a chromogenic analog of lactose, truns blue when B-galactosidase is present, and remains white in its absence, so blue-white screening can indicate which colonies contain recombinant plasmids.
Blue white screen • A portion of the lactose utilizing gene (b-galactosidase) is interrupted by the polylinker cloning site. • Insertion of a DNA fragment prevents expression of the gene. • Growing the E. coli containing the plasmid on petri plates containing a substrate for the enzyme allows you to tell those which express the lac-z gene (no insert, blue color) form those colonies that do not (contain an insert, white color)
How to insert the DNA? • First digest then ligate: • Cut pUC19 with an RE that has a unique site in the polylinker. • Cut the DNA to be cloned (insert DNA) with the same enzyme. • Mix insert DNA and pUC19 DNA • Transform plasmids into E. coli, either through chemical treatment of cells or electroporation. • Incubate the recombinant DNA plasmids with E.coli cells treated chemically to take up DNA • Electroporation requires that an electric shock is delivered to the cells causing temporary disruptions of the cell membrane and letting the DNA enter.
How to insert the DNA? • Grow cells on media containing ampicillin and X-gal. • Ampicillin-resistant colonies will grow. • Blue colonies contain only the vector • White colonies contain vector with insert.
Other plasmid vectors • They contain • different unique restriction enzyme sites • Phage promoters (e.g., T7, T3, SP6) for transcription of the cloned DNA. • Available for prokaryotic and eukaryotic organisms • Size of the insert is limited; plasmids carrying more than 5-10 kb are not stable.
Phage Lambda Vectors • Versions of bacteriophage lambda with sequences for lysogeny removed, so that only lytic infection is possible. • Lambda replacement vector: • Has a chromosome with a ‘left’ arm and a ‘right’ arm, that contain all the genes needed for lysis. Between two arms, there is a disposable segment since it does not contain any lytic cycle genes. These two regions, the arms and the disposable region is separated by EcoRI sites. The lambda chromosome central region is replaced with the insert DNA (~15kb), using RE digestion and ligation.
Phage l (lambda) as a Cloning Vector • Plasmids are limited in the size of DNA that can be easily introduced into bacteria, about 5-10 Kb cloned DNA (transformation). By cloning into a phage, the viral entry system can be exploited to introduce the DNA into bacteria. Phage l allows insertion of 15-30 Kb DNA, with efficient introduction into E. coli. • Subcloning: transfer of a DNA insert from the phage clone into a plasmid by having a special bacterial strain do the work.
Only phages with DNA insert between the two arms can replicate; Why? • The phage needs both arms to be together for reproduction and lysis. • Each DNA fragment is cloned by repeated rounds of infection and lysis. Eventually culture becomes transparent as all the bacteria has been lysed, and a population of progeny lambda phages is produced (1010 to 1011 phages/mL).
Phage vectors • When ligated DNA is mixed with phage lambda proteins, phage heads assemble, and DNA is packaged, forming virus particles. • Only phages with both arms of the phage lambda chromosome and a properly sized (37-52 kb) central insert sequence are able to replicate by infecting E. coli.
Shuttle Vectors • A cloning vector capable of replicating in two or more types of organism (e.g., E.coli and yeast) is called a shuttle vector. They replicate autonomously in both hosts or integrate into them.
Yeast Artificial Chromosomes (YACs) • Function as artificial chromosomes in yeast. • Linear structure with a yeast telomere (TEL) at each end. • A yeast centromere sequence (CEN) • A marker gene on each arm that is selectable in yeast (e.g., TRP1, URA3). • Tryptophan and uracil independence in trp1 and ura3 mutant strains, respectively. • Unique restriction sites for inserting foreign DNA that can be up to 500kb long. This size of inserts are important in generating physical maps of genomes. • An origin of replication sequence-ARS (autonomously replicating sequence)-that allows the vector to replicate in yeast.
YACs • Several hundred kb of insert DNA can be cloned in a YAC. YAC clones are made by: • Generating YAC arms by restriction digest • Ligating with insert fragments up to 500 kb in length • Transforming into yeast • Selecting for markers (e.g., TRP1 and URA3).
Question 7.8 • Why one might want to clone DNA in an organism other than E.coli. • One can transform yeast as well as plant and animal cells. This can be useful in studying cloned eukaryotic genes in a eukaryotic environment, commercial production of gene products (e.g., drugs, antibodies), developing gene therapy, genetic engineering.
Question 7.8 • Why use shuttle vectors? • They can replicate in two or more host systems, e.g., replicate in E.coli (it is easy to do the initial cloning), then be transferred to yeast. A yeast shuttle vector contains selectable markers for both systems (ura3 for yeast; ampR for E. coli); autonomous replication sites to replicate as a plasmid.
Recombinant DNA libraries • Genomic library • Chromosome library • cDNA library
Genomic libraries • Are constructed by digesting genomic DNA • Complete digestion • Mechanical shearing • Partial digestion
Partial digestion • They are selected in a certain size range by density gradient centrifugation or agarose gel electrophoresis • DNA fragments from sticky ends can be cloned directly. • Genomic sequences are not equally represented in the library • Regions of DNA with relevant restriction sites very close together or far apart are removed at the selection stage • Some regions prevent vector replication so eliminated.
Partial Digest for Producing Clonable Fragments Enzymes with compatible Sticky ends are used.
How many clones are needed to contain all sequences in genome? • Depends on: • Size of the genome being cloned • The average size of the DNA fragments inserted into the vector.
How many clones are needed to contain all sequences in genome? • The probability of having at least one copy of any DNA sequence in the genomic library is: • N=ln(l-P)/ln(1-f), • Where N = the necessary number of recombinant DNA molecules; • P = the probability desired; • F = average size of the fragments divided by the genome size • Ln = natural algorithm
Question 7.10 • Within the human genome (3 x 109 bp), how many 40-kb pieces would you have to clone into a library if you wanted to be 90% certain of including a particular sequence? • P = 0.90; f=40,000/(3x109) • N=ln(l-P)/ln(1-f) = 172,693 fragments
cDNA libraries • cDNA drives from mature mRNA, no introns. • polyA tail at the 3’ is useful for: • Isolating mRNA from cell lysates • Priming the synthesis of cDNA providing a known 5’ sequence
cDNA synthesis • A short oligo(dT) primer is used. It anneals to the mRNA’s poly(A) tail, allows reverse transcriptase to synthesize the cDNA (DNA-mRNA hybrid). • Rnase H degrades the mRNA strand, creating small fragments that serve as primers • DNA polymerase I makes new DNA fragments, DNA ligase connects them to make a complete chain.
cDNA from a Polyadenylated mRNA Annealing Reverse transcription RnaseH degradation DNA polymerase I DNA ligase
cDNA cloning • Introduction of restriction site linkers to the ends of the cDNA by blunt end ligation • Digestion with cognate restriction enzyme to create sticky ends • Mixing cDNA with vector DNA cut with the same restriction enzyme in the presence of DNA ligase • Transforming into an E. coli host for cloning • Use polylinkers engineered with appropriate ssDNA overhangs so do not digest
Linkers for Cloning DNA • Any DNA fragment can have a specific restriction site added to the ends by ligating on a “linker”. • Linkers are small, synthetic (made in the lab, or ordered from a company) DNA fragments which contain the recognition sequence for one or more restriction enzymes. • After ligating on linkers, the DNA is cut with the appropriate restriction enzyme to produce ends for cloning.
Random Primed DNA Synthesis for Making a “Probe” • One common technique for making a probe to detect a specific DNA sequence is called “random primed DNA synthesis”. • Short (8 bases) primers of random sequence are annealed to the heat denatured DNA (of the sequence for the probe), and DNA polymerase is used to synthesize copies of the DNA with one of the nucleotides incorporated carrying a detectable label (such as radioactive phosphate).
Klenow fragment PolI • The large or Klenow fragment of DNA polymerase I has DNA polymerase and 3' -> 5' exonuclease activities, and is widely used in molecular biology.
cDNA synthesis & making probe • DNA polymerases require a primer (oligonucleotides, 6-20 bases) to provide a free 3' hydroxyl group for initiation of synthesis. Mixed single-stranded template (usually denatured double-standed DNA), primers and the enzyme in the presence of an appropriate buffer. As Klenow proceeds, it can displace primers downstream and continue synthesizing new DNA.
Colony Lift Hybridization to Find a Cloned Sequence in a Plasmid (or Cosmid) Library • The presence of a clone containing a specific sequence can be determined by making a “lift” of the colonies, lysing the cells on the surface of a “membrane”, and hybridizing a labeled (radioactive) “probe” of the sequence being searched for.
Finding a Cloned Sequence: Plaque Lift Hybridization in a Lambda Library • If a library is made in phage l, the desired sequence can be found by hybridizing a probe to a “lift” of the plaques. This is detected with a probe, as in the colony lift.