Download
1 / 50
510 likes | 892 Views
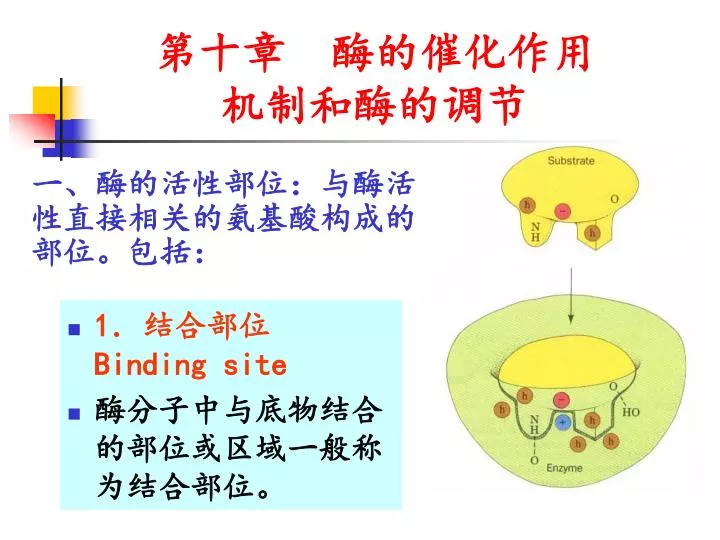
第十章 酶的 催化作用 机制和酶的调节. 1 .结合部位 Binding site 酶分子中与底物结合的部位或区域一般称为结合部位。. 一、酶的活性部位:与酶活性直接相关的氨基酸构成的部位。包括:. 2 .催化部位 catalytic site. 酶分子中促使底物发生化学变化的部位称为催化部位 。 通常将酶的结合部位和催化部位总称为酶的 活性部位或活性中心 。 结合部位决定酶的专一性, 催化部位决定酶所催化反应的性质。. 补:酶活性中心的必需基团 顺便复习一些重要 AA 的 R 基. 酶必需基团 指酶活性中心实现催化作用的必需的些氨基酸基团。 主要包括:

E N D
第十章 酶的催化作用机制和酶的调节 • 1.结合部位 Binding site • 酶分子中与底物结合的部位或区域一般称为结合部位。 一、酶的活性部位:与酶活性直接相关的氨基酸构成的部位。包括:
2.催化部位 catalytic site • 酶分子中促使底物发生化学变化的部位称为催化部位。 • 通常将酶的结合部位和催化部位总称为酶的活性部位或活性中心。 • 结合部位决定酶的专一性, • 催化部位决定酶所催化反应的性质。
补:酶活性中心的必需基团 顺便复习一些重要AA的R基 • 酶必需基团指酶活性中心实现催化作用的必需的些氨基酸基团。 • 主要包括: • 亲核性基团:丝氨酸的羟基,半胱氨酸的巯基和组氨酸的咪唑基。
酸、碱性基团:天冬氨酸和谷氨酸的羧基,酪氨酸的酚羟基酸、碱性基团:天冬氨酸和谷氨酸的羧基,酪氨酸的酚羟基 • 赖氨酸的氨基,组氨酸的咪唑基。
补:酶的调节部位 Regulatory site • 某些酶分子中存在着一些可以与其他分子发生某种程度的结合的部位,从而引起酶分子空间构象的变化,对酶起激活或抑制作用。
酶活性部位的共同特点 • (1)活性部位酶占总体很小部分:1-2% • (2)酶活性部位是一个三维实体 • (3)酶活性部位与底物通过诱导契合后结合 • (4)酶活性部位是其表面一个裂缝,内部非极性基团较多,是一疏水区域 • (5)底物通过非共价与酶结合 • (6)活性部位具有柔性或可运动性
著名生物化学家、中科院院士——邹承鲁 • 邹承鲁(1923年5月17日—),出生于山东青岛,祖籍江苏无锡。生物化学家。1980年当选为中国科学院学部委员(院士)。 • 1946年,赴英国剑桥大学。研究生期间,他在国际上最早用蛋白水解酶部分水解方法研究蛋白质结构与功能的关系,单独署名的论文在英国《自然》杂志发表。1951年,邹承鲁获英国剑桥大学生物化学博士学位。 • 1958年,他参加发起人工合成胰岛素工作,并负责胰岛素A和B链的拆合。 • 邹承鲁等用自己创立的动力学方法比较研究了各种不同类型的酶在变性过程中构象和活力变化的关系,得到活性丧失先于构象变化的结果,据此提出了酶活性部位柔性的假说。 • 邹承鲁:我的学生差不多都退休了。 • 学生:就先生还在这干。我们也快退休了,但是老先生还得干。 • 邹承鲁:我是退休嫌闷得慌。 • 学生:其实搞科学的就是这样,他也不求什么,就求一个能干自己想干的事的环境。
酶催化作用的本质 • 酶催化作用的本质是酶的活性中心与底物分子通过短程非共价力(如氢键,离子键和疏水键等)的作用,形成E-S反应中间物。 • 其结果使底物的价键状态发生形变或极化,起到激活底物分子和降低过渡态活化能作用。
*二、酶催化反应的独特性质 (自学,以增对酶催化作用所理解) 三、影响酶催化效率的有关因素 • 酶是专一性强,催化效率很高的生物催化剂,这是由酶分子的特殊结构决定的。多种因素可以使酶催化的反应加速:
1、底物和酶的邻近效应和定向效应 • 邻近效应:酶与底物形成复合物后,使底物和底物之间(对多底物反应而言),酶的催化基团与底物之间结合于同一分子使反应有效浓度大提高,有利于提高反应速率大增加。 • 定向效应:反应物的反应基团之间和酶的催化基团与底物的反应基团之间正确取位,使底物分子中参与反应的基团相互接近,并被严格定定位,使酶促反应具有高效率和专一性特点。(例如:P389邻苯丙酸内酯形成的反应)
例:邻近效应 以乙酸对硝基苯酯以咪唑作为酶催化的水解反应为例,若将咪唑连到其上,则反应加快24倍
2、底物形变和诱导契合 • 该学说认为酶表面并没有一种与底物互补的固定形状,而只是由于底物的诱导才形成了互补形状,同时底物了发生形变,接近其过渡态,降低了反应活化能,使反应易于发生. • 形变的机制:与酶结合,底物分子内敏感键中电子云密度增高或降低,使敏感键更敏感,从而底物分子形变。
3、 酸碱催化 • 指通过瞬时的向反应物提供质子或从反应物接受质子以稳定过渡态,加速反应的催化机制。 • 酸-碱催化可分为狭义的酸-碱催化和广义的酸-碱催化。酶参与的酸-碱催化反应一般都是广义的酸-碱催化方式。 • 广义酸-碱催化是指通过质子酸提供部分质子,或是通过质子碱接受部分质子的作用,达到降低反应活化能的过程。
酶分子中可以作为广义酸、碱的基团 • 广义酸基团 广义碱基团(质子供体) (质子受体) His 是酶的酸碱催化作用中最活泼的一个催化功能团。
4、 共价催化 • 基本原理: • 又称亲核催化或亲电子催化,催化时,酶分子上亲核基团或亲电子基团分别放出电子或汲取电子并作用于底物的缺电子或富电子中心,迅速形成不稳定的共价中间复合物,降低反应活化能,使反应加速。 • 酶上常见的亲核基团: Ser 的羟基, Cys 的巯基, His 的咪唑基。 • 底物上亲电基团:磷酰基,酰基,糖基。——P392实例
5、 金属离子催化作用 *根据金属离子与酶蛋白结合程度,可分为两类:金属酶和金属—激活酶。 (1)金属酶:酶蛋白与金属离子结合紧密。如 Fe2+/ Fe3+ 、Cu+/Cu3、Zn2+ 、Mn2+、Co2等过渡态离子。 (2)金属—激活酶:金属离子与酶的结合一般较松散。在溶液中,酶与这类离子结合而被激活。如Na+ 、K+、 Mg2+、 Ca2+等。 以3种主要途径参加催化过程: (1)通过结合底物为反应定向
(2)通过可逆改变金属离子氧化态调节氧化还原反应(2)通过可逆改变金属离子氧化态调节氧化还原反应 • 许多氧化-还原酶中都含有铜或铁离子,它们作为酶的辅助因子起着传递电子的功能。 • Fe3+e Fe2+ • Cu2+e Cu+
(3)通过静电稳定或屏蔽负电荷 • 电荷屏蔽作用是酶中金属离子的一个重要功能。 • 如:Mg2+-ATP复合物。
6、 多元催化和协同效应 • 酶的活性中心部位,一般都含有多个起催化作用的基团,这些基团配合在一起作用如胰凝乳蛋白酶通过Asp102、His57和Ser195组成的电荷中继网催化肽键水解。
7、 活性部位微环境影响 • 酶的活性中心部位处于疏水环境裂缝中,是一个非极性环境,其介电常数较低,从而使处于其中带电基团之间的静电作用比在极性环境中显著增高,因此有利于催化反应。
四、酶催化反应机制的实例 • *溶菌酶, *胰核糖核酸酶A,*羧肽酶A,丝氨酸蛋白酶族,天冬氨酸蛋白酶 • (一)溶菌酶 • 存在于蛋清及眼泪,催化某些细菌胞壁多糖(N-乙酰葡糖胺[NAG]+N-乙酰胞壁酸[NAM])水解,从而抗菌 • 14.6kD,129个AA,单肽链,4对二硫键,表而有一裂缝,能容纳多糖底物的6个单糖,为酶的活性部位。P395图10-9
(二)胰核糖核酸酶A • 124个AA,单肽链,4对二硫键 • 水解RNA,专一切开3’P-O键,生成嘧啶核苷3’-磷酸或以其结尾的寡聚核苷酸 • His119和His12为酶活性中心必需基团(见下图)
碱催化 酸催化
(三)羧肽酶307个AA,单肽链,紧密结合一个Zn2+催化肽链C末端肽键水解(三)羧肽酶307个AA,单肽链,紧密结合一个Zn2+催化肽链C末端肽键水解 锌离子在酶活性部位 与2His和Glu配位连接
底物(Gly-Tyr)与酶诱导契合进行结合P403-404 由145,248,270位 的AA和底物结合,将 进一步由锌离子,Arg127,Glu270对底物催化
(四)丝氨酸蛋白酶族 • 包括:胰蛋白酶,胰凝乳蛋白酶,弹性蛋白酶等 • 三种酶分子量范围在2.5kD,具有相似顺序和三级结构
三种酶的底物结合口袋 非常保守
三、酶活性的调节控制 • (一)别构调控 • 1、别构调节:酶分子的非催化部位与某些化合物可逆地非共价结合后发生构象的改变,进而改变酶活性状态的作用。 • 2、别构酶:具有上述这种调节作用的酶。 • 3、效应物:能使酶分子发生别构作用的物质。可分为正效应物(别构激活剂)和负效应物(别构抑制剂)。 • *ATCase, 3-磷酸甘油醛脱氢酶
1、天冬氨酸转氨甲酰酶(ATCase) (1)合成途径终产物CTP对该酶有反馈抑制作用,ATP与 CTP竞争调节部位,可阻止CTP的抑制作用——异促效应
(2)含2个催化亚基[3条肽链]和3个调节亚基[2条肽链],催化亚基无与ATP或CTP结合能力;调节亚基可结合ATP或CTP,无催化能力。调节亚基与ATP或CTP结合为非共价。(2)含2个催化亚基[3条肽链]和3个调节亚基[2条肽链],催化亚基无与ATP或CTP结合能力;调节亚基可结合ATP或CTP,无催化能力。调节亚基与ATP或CTP结合为非共价。
3、别构酶的性质 • (1)一般都是寡聚酶,通过次级键由多亚基构成。 • (2)别构酶动力学曲线不是双曲线,不符合米氏方程,正协同效应为S形典线,负协同效应为表观双曲线。 • 补(3)酶上含催化亚基和调节亚基,效应物和调节亚基以非共价结合,改变酶的构象,影响其与底物结合,从而调节催化反应。
1、非别构酶 2、正协同效应 别构酶 3、负协同效应别构酶
4、别构模型 • (1)协同模型(WMC模型):又称齐变模型,即在酶活性调节中,当蛋白质构象变化时各亚基几乎同时从一种构象转变为另一种构象。 • (2)序变模型(KNF模型):各亚基的构象以序变方式进行。 • 分别见后图
(二)酶原的激活 • 1、酶原的激活:体内合成的蛋白质,有时不具有生物活性,经过蛋白水解酶专一作用后,构象发生变化,形成酶的活性部位,变成具有活性的酶的过程。这个不具生物活性的蛋白质称为前体。若活性蛋白是酶,该前体称为酶原。 • 2、举例:胰凝乳蛋白酶原的激活,胰蛋白酶的激活,胃蛋白酶的激活
肠激酶 -胰凝乳蛋白酶活性最高,但不稳定 -胰凝乳蛋白酶稳定,活性只有的40% -
思考 • 机体为会么先产生酶原后再经过剪切加工形成活性酶,为会么不直接产生活性酶,其生理意义何在?
(三)可逆的共价修饰 • 1、共价调节酶:可被共价修饰而改变酶活性。 • 2、可逆的共价修饰:通过其它酶对其多肽链上某些基团进行可逆的共价修饰,使酶处于活性与非活性的互变状态,从而调节酶活性。 • 3、例如:蛋白激酶通过蛋白磷酸化和去磷酸化来失现酶活性与非活性的互变。
六、同工酶 • 1、同工酶:指能催化相同化学反应,但其蛋白质分子结构、理化性质和免疫性能等方面都存在明显差异的一组酶。 • 2、存在:(1)同一个体的不同组织中; • (2)同一组织、同一细胞的不 • 同亚细胞结构中。 • 3、例如:LDH(乳酸脱氢酶)有5种同工酶:LDH1(H4),LDH2(MH3),LDH3(M2H2),LDH4(M3H),LDH5(M4) • 4、同工酶的作用或功能:见书428-429页。
习题 活性中心和必需基团 酶原 同工酶巯基酶酶原激活金属酶 别构效应剂诱导酶(补)协同效应KNF模型别构酶 共价修饰调节酶的反馈抑制MWC模型 酶原 在效应物作用下,蛋白质产生的变构(或别构)效应是蛋白质的 A 一级结构发生变化 B 构型发生变化 C 构象发生变化 D 氨基酸顺序发生变化 在嘧啶核苷酸的合成途径中,CTP可以使天冬氨酸转氨甲酰酶产生别构效应的事实属于下列哪种情况: A. 别构抑制 B. 别构激活 C.酶的诱导生成作用 D. 非共价作用 E. 前体活化作用 关于变构酶的结构特点的错误叙述是: A、有多个亚基组成 B、有与底物结合的部位 C、有与变构剂结合的部位 D、催化部位与别构部位都处于同一亚基上 E、催化部位与别构部位既可处于同一亚基也可处于不同亚基上
习题 * 关于别构酶,正确的表达是 A、它们一般是寡聚酶 B、它们一般是单体酶 C、当效应剂与别构酶非共价结合后,引起别构效应。 D、当效应剂与别构酶共价结合后,引起别构效应。 E、别构酶的动力学性质符合米氏方程式。 F、别构酶的动力学性质不符合米氏方程式。 下列哪一个酶的催化活性需要金属离子? A、溶菌酶B、羧肽酶C、胰凝乳蛋白酶D、胰蛋白酶 胰蛋白酶属单体酶。 别构酶动力学曲线的特点都是呈S形曲线。 酶活性中心是亲水的介电区域。 溶菌酶的实现催化功能需金属离子Mg2+参与。 4、 简述酶活性调控的几种机制。 同工酶有何生理意义?



