Download
1 / 1
10 likes | 292 Views
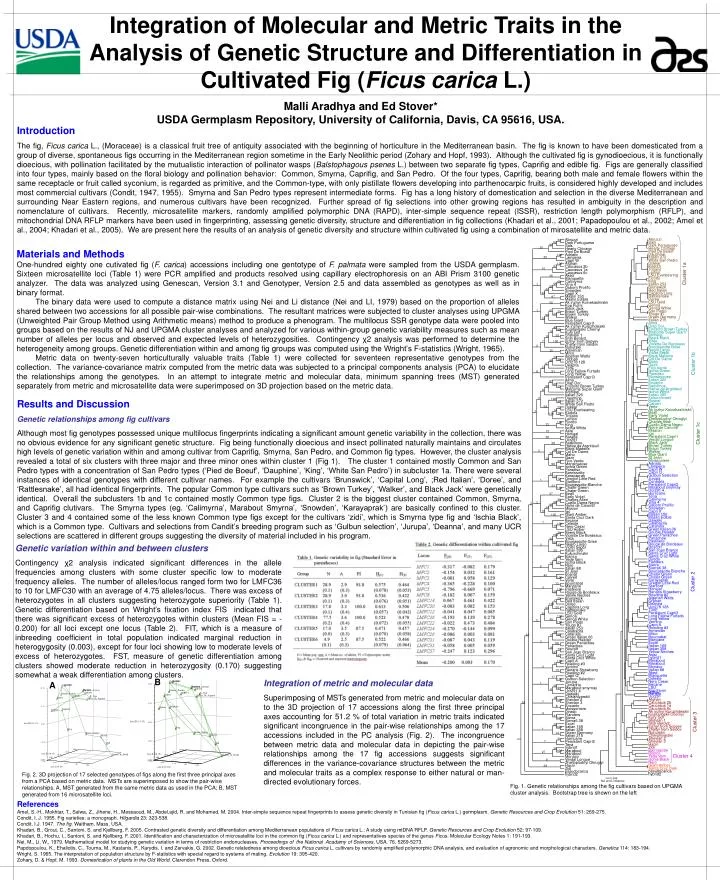
B. A. Abruzzi. 34. 62. Dark Portuguese. 99. Sals. 30. Hearty Chicago. Pied De Boeuf. Abruzzi. Adriatic. 100. Sals. Lampeira. 30. Dark Portuguese. Capri W. Hearty Chicago. Afghan. 100. Pied De Boeuf. 27. Caucasus 2b. Dauphine. 10. Caucasus 1a. Italian 372. Caucasus 6c.

E N D
B A Abruzzi 34 62 Dark Portuguese 99 Sals 30 Hearty Chicago Pied De Boeuf Abruzzi Adriatic 100 Sals Lampeira 30 Dark Portuguese Capri W Hearty Chicago Afghan 100 Pied De Boeuf 27 Caucasus 2b Dauphine 10 Caucasus 1a Italian 372 Caucasus 6c White San Pedro Aked Dokkar 26 Blanquette Kadota Trojano Calimyrna 65 Lemon Cluster 1a 55 Vina 4 1 LSU Everbearing 55 Osborn Prolific Ficotto 40 Snowden King 22 Capri X Italian 253 Maslin 150 97 Brunswick Maslin Edible Red Italian Ak inzhyr Koinekashirskii 20 Capitola Long Kop Fiomi Rattlesnake Black Jack Doree 1 91 Braun Turkey 31 LSU Gold Brown Turkey 22 Genoa 99 Walker Genoa White 10 Blue Giant San Pietro Persistent Capri1 Yougo #7 Ak inzhyr Kuruzhdeiskii Green Germany 49 Italian 215 Kugitangskii Chernyi 98 15 Alma Kury Gol Drap Dor Shevlan 1 29 10 Encanto Brown Turkey Shih Berdy2 Malcoms Super Giant Inzhyr from Sopyev 7 100 Archipel Zheltyi from Seidov Beers Black 2 Nuhurskii Vista Armenian 20 Violette De Bordeaux Milco Bourjassotte Grise Algerian Watts Negro Largo 24 Orphan 28 Violet Sepor 98 Ucr278 128 Black Madeira 67 Deanna Col De Dame 25 T30E Maho Cluster 1b Long Fellow Furtado Gazir 91 6 Long Yellow Fico Verde Persistent Capri3 Ischia Green Paradiso Alma 87 Monstrueuse Drap Dor 60 Italian 320 Encanto Brown Turkey 96 33 Encanto Malcoms Super Giant 8 Barbillone Archipel Hative de Argntteuil Italian 320 Ischia White Dauphine 3 100 Italian 395 Italian 372 36 Kukurchinskii White San Pedro Bosnat 64 Dokkar 22 Calvert LSU Everbearing 14 Verte 1 Kadota 19 Ak inzhyr Koinekashirskii 100 Trojano Beall Lemon Early Violet Ficotto Zheltoplodnyi Okruglyi 100 23 King Charles Allen Ischia White Cuello Dama Negro Noire de Caromb Asisi 13 Mission Honigal Cluster 1c Igo Barada 28 Persistent Capri1 Ucr291 Verdal Longue Barbillone 38 Black Jack Hative de Argntteuil Brown Turkey Black Madeira 35 Braun Turkey 3 Col De Dame 98 Walker 22 Maho Blue Giant Gazir 16 St Jean Fico Verde 34 Moissoniere Monstrueuse 99 Adriatic Ischia Green Lampeira 28 Paradiso Capri W Barnissotte Capri Q 3 Karayaprak Gulbun Selection 20 Oregon Little Red Jurupa 72 1 Stanford Conadria Persistent Capri2 Bourjassotte Blanche 23 Marabout smyrnay Chater Green 100 Ucr291 4 Chater Green Hurricane Beall 37 Tena Early Violet Calimyrna Charles Allen 36 Vina 4 68 Cuello Dama Negro 2 Osborn Prolific 2 96 Noire de Caromb Snowden 34 Mission Capri X Igo 10 Maslin 150 Giant Amber 100 Maslin Edible Santa Cruz Dark Dawalki Becane Calabacita 7 Celeste Catarulla 100 Nero Cesar 15 Green Italian 06 LSU Hollier Double Header Beers Black Green Panachee 35 Panachee 100 Violette De Bordeaux Pastiliere Vista 34 Rouge de Bordeaux Bourjassotte Grise 99 Roscoff Negro Largo 3 43 San Joao Branco Violet Sepor Santa Cruz Light Italian 395 25 Santa Cruz White Kukurchinskii Diredo Bianco 45 Flanders Yede Vern 22 Sierra Ischia Black 27 Sierra6 38 12 Nero Bourjassotte Blanche 6 Italian 88 Chater Green St Jean Chater Green Bosnat Karayaprak Cluster 2 40 Calvert Oregon Little Red 100 Verte Stanford Bournabat Honigal 13 Renans Strawberry Marylane 3 Roeding #2 Pastiliere 100 Algerian Watts 26 Rouge de Bordeaux Orphan Yellow Neches Deanna Brunswick 20 Ucr278 128 Red Italian 17 T30E Doree 100 Persistent Capri3 Capitola Long 20 63 Long Fellow Furtado Rattlesnake Long Yellow LSU Gold 30 Vernino Genoa 100 Capri A Genoa White Roeding #3 65 2 San Pietro Armenian 100 Yougo #7 Milco Italian 253 Bournabat Calabacita Marylane 39 Excel Catarulla 96 42 11 Italian 169 Green Italian 06 Italian 358 Double Header 3 Yellow Neches Green Panachee 99 Asisi Panachee hybrid1 16 Roscoff 73 Marabout San Joao Branco 93 Marabout Santa Cruz Light 47 Monaco Santa Cruz White Italian 88 Capri A 100 Aked Roeding #3 9 Blanquette Vernino 2 Celeste Renans Strawberry Nero Cesar 13 Roeding #2 Becane Capri Q LSU Hollier 17 Gulbun Selection Kop Fiomi 100 Jurupa Barada 1 Ucr291 Conadria 26 16 Afghan Marabout smyrnay 100 91 4 7 Caucasus 2b Ucr291 4 Caucasus 1a Dawalki 8 Caucasus 6c Chikishlyarskii Ak inzhyr Kuruzhdeiskii Shevlan 2 4 Kugitangskii Chernyi Shevlan 3 52 41 Kury Gol 100 Encanto Cluster 3 Shevlan 1 Moissoniere Shih Berdy2 Diredo 29 Inzhyr from Sopyev 100 Flanders 1 Zheltyi from Seidov Sierra Nuhurskii 14 Sierra6 38 Chikishlyarskii Excel Shevlan 2 31 Italian 169 Shevlan 3 7 Italian 358 Hacin Green Germany Zidi 34 Italian 215 Barnissotte 100 27 Bianco Hurricane Cluster 4 Yede Vern Persistent Capri2 Ischia Black Tena 8 Nero hybrid1 Giant Amber Marabout 28 Santa Cruz Dark Marabout pseudocarica Monaco hybrid2 Verdal Longue Zheltoplodnyi Okruglyi 0.005 Nei and Li distance Hacin Zidi pseudocarica hybrid2 Integration of Molecular and Metric Traits in the Analysis of Genetic Structure and Differentiation in Cultivated Fig (Ficus carica L.) Malli Aradhya and Ed Stover* USDA Germplasm Repository, University of California, Davis, CA 95616, USA. Introduction The fig, Ficus carica L., (Moraceae) is a classical fruit tree of antiquity associated with the beginning of horticulture in the Mediterranean basin. The fig is known to have been domesticated from a group of diverse, spontaneous figs occurring in the Mediterranean region sometime in the Early Neolithic period (Zohary and Hopf, 1993). Although the cultivated fig is gynodioecious, it is functionally dioecious, with pollination facilitated by the mutualistic interaction of pollinator wasps (Balstophagous psenes L.) between two separate fig types, Caprifig and edible fig. Figs are generally classified into four types, mainly based on the floral biology and pollination behavior: Common, Smyrna, Caprifig, and San Pedro. Of the four types, Caprifig, bearing both male and female flowers within the same receptacle or fruit called syconium, is regarded as primitive, and the Common-type, with only pistillate flowers developing into parthenocarpic fruits, is considered highly developed and includes most commercial cultivars (Condit, 1947, 1955). Smyrna and San Pedro types represent intermediate forms. Fig has a long history of domestication and selection in the diverse Mediterranean and surrounding Near Eastern regions, and numerous cultivars have been recognized. Further spread of fig selections into other growing regions has resulted in ambiguity in the description and nomenclature of cultivars. Recently, microsatellite markers, randomly amplified polymorphic DNA (RAPD), inter-simple sequence repeat (ISSR), restriction length polymorphism (RFLP), and mitochondrial DNA RFLP markers have been used in fingerprinting, assessing genetic diversity, structure and differentiation in fig collections (Khadari et al., 2001; Papadopoulou et al., 2002; Amel et al., 2004; Khadari et al., 2005). We are present here the results of an analysis of genetic diversity and structure within cultivated fig using a combination of mirosatellite and metric data. Materials and Methods One-hundred eighty one cutivated fig (F. carica) accessions including one gentotype of F. palmata were sampled from the USDA germplasm. Sixteen microsatellite loci (Table 1) were PCR amplified and products resolved using capillary electrophoresis on an ABI Prism 3100 genetic analyzer. The data was analyzed using Genescan, Version 3.1 and Genotyper, Version 2.5 and data assembled as genotypes as well as in binary format. The binary data were used to compute a distance matrix using Nei and Li distance (Nei and LI, 1979) based on the proportion of alleles shared between two accessions for all possible pair-wise combinations. The resultant matrices were subjected to cluster analyses using UPGMA (Unweighted Pair Group Method using Arithmetic means) method to produce a phenogram. The multilocus SSR genotype data were pooled into groups based on the results of NJ and UPGMA cluster analyses and analyzed for various within-group genetic variability measures such as mean number of alleles per locus and observed and expected levels of heterozygosities. Contingency χ2 analysis was performed to determine the heterogeneity among groups. Genetic differentiation within and among fig groups was computed using the Wright’s F-statistics (Wright, 1965). Metric data on twenty-seven horticulturally valuable traits (Table 1) were collected for seventeen representative genotypes from the collection. The variance-covariance matrix computed from the metric data was subjected to a principal components analysis (PCA) to elucidate the relationships among the genotypes. In an attempt to integrate metric and molecular data, minimum spanning trees (MST) generated separately from metric and microsatellite data were superimposed on 3D projection based on the metric data. Results and Discussion Genetic relationships among fig cultivars Although most fig genotypes possessed unique multilocus fingerprints indicating a significant amount genetic variability in the collection, there was no obvious evidence for any significant genetic structure. Fig being functionally dioecious and insect pollinated naturally maintains and circulates high levels of genetic variation within and among cultivar from Caprifig, Smyrna, San Pedro, and Common fig types. However, the cluster analysis revealed a total of six clusters with three major and three minor ones within cluster 1 (Fig 1). The cluster 1 contained mostly Common and San Pedro types with a concentration of San Pedro types (‘Pied de Boeuf’, ‘Dauphine’, ‘King’, ‘White San Pedro’) in subcluster 1a. There were several instances of identical genotypes with different cultivar names. For example the cultivars ‘Brunswick’, ‘Capital Long’, ;Red Italian’, ‘Doree’, and ‘Rattlesnake’, all had identical fingerprints. The popular Common type cultivars such as ‘Brown Turkey’, ‘Walker’, and Black Jack’ were genetically identical. Overall the subclusters 1b and 1c contained mostly Common type figs. Cluster 2 is the biggest cluster contained Common, Smyrna, and Caprifig clutivars. The Smyrna types (eg. ‘Calimyrna’, Marabout Smyrna’, ‘Snowden’, ‘Karayaprak’) are basically confined to this cluster. Cluster 3 and 4 contained some of the less known Common type figs except for the cultivars ‘zidi’, which is Smyrna type fig and ‘Ischia Black’, which is a Common type. Cultivars and selections from Candit’s breeding program such as ‘Gulbun selection’, ‘Jurupa’, ‘Deanna’, and many UCR selections are scattered in different groups suggesting the diversity of material included in his program. Genetic variation within and between clusters Contingency χ2 analysis indicated significant differences in the allele frequencies among clusters with some cluster specific low to moderate frequency alleles. The number of alleles/locus ranged form two for LMFC36 to 10 for LMFC30 with an average of 4.75 alleles/locus. There was excess of heterozygotes in all clusters suggesting heterozygote superiority (Table 1). Genetic differentiation based on Wright’s fixation index FIS indicated that there was significant excess of heterozygotes within clusters (Mean FIS = -0.200) for all loci except one locus (Table 2). FIT, which is a measure of inbreeding coefficient in total population indicated marginal reduction in heterogygosity (0.003), except for four loci showing low to moderate levels of excess of heterozygotes. FST, measure of genetic differentiation among clusters showed moderate reduction in heterozygosity (0.170) suggesting somewhat a weak differentiation among clusters. Integration of metric and molecular data Superimposing of MSTs generated from metric and molecular data on to the 3D projection of 17 accessions along the first three principal axes accounting for 51.2 % of total variation in metric traits indicated significant incongruence in the pair-wise relationships among the 17 accessions included in the PC analysis (Fig. 2). The incongruence between metric data and molecular data in depicting the pair-wise relationships among the 17 fig accessions suggests significant differences in the variance-covariance structures between the metric and molecular traits as a complex response to either natural or man-directed evolutionary forces. Fig. 2. 3D projection of 17 selected genotypes of figs along the first three principal axes from a PCA based on metric data. MSTs are superimposed to show the pair-wise relationships. A, MST generated from the same metric data as used in the PCA; B, MST generated from 16 microsatellite loci. Fig. 1. Genetic relationships among the fig cultivars based on UPGMA cluster analysis. Bootstrap tree is shown on the left References Amel, S.-H., Mokhtar, T., Salwa, Z., Jihene, H., Messaoud, M., Abdel,ajid, R. and Mohamed, M. 2004. Inter-simple sequence repeat fingerprints to assess genetic diversity in Tunisian fig (Ficus carica L.) germplasm. Genetic Resources and Crop Evolution 51: 269-275. Condit, I. J. 1955. Fig varieties: a monograph. Hilgardia 23: 323-538. Condit, I.J. 1947. The fig. Waltham, Mass, USA. Khadari, B., Grout, C., Santoni, S. and Kjellberg, F. 2005. Contrasted genetic diversity and differentiation among Mediterranean populations of Ficus carica L.: A study using mtDNA RFLP. Genetic Resources and Crop Evolution 52: 97-109. Khadari, B., Hochu, I., Santoni, S. and Kjellberg, F. 2001. Identification and characterization of microsatellite loci in the common fig (Ficus carica L.) and representatives species of the genus Ficus. Molecular Ecology Notes 1: 191-193. Nei, M., Li, W., 1979. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proceedings of the National Acadamy of Sciences, USA. 76, 5269-5273. Papdopoulou, K., Ehaliotis, C., Tourna, M., Kastanis, P., Karydis, I. and Zervakis, G. 2002. Genetic relatedness among dioecious Ficus carica L. cultivars by randomly amplified polymorphic DNA analysis, and evaluation of agronomic and morphological characters. Genetica 114: 183-194. Wright, S. 1965. The interpretation of population structure by F-statistics with special regard to systems of mating. Evolution 19: 395-420. Zohary, D. & Hopf, M. 1993. Domestication of plants in the Old World. Clarendon Press, Oxford.