Download
1 / 28
290 likes | 467 Views
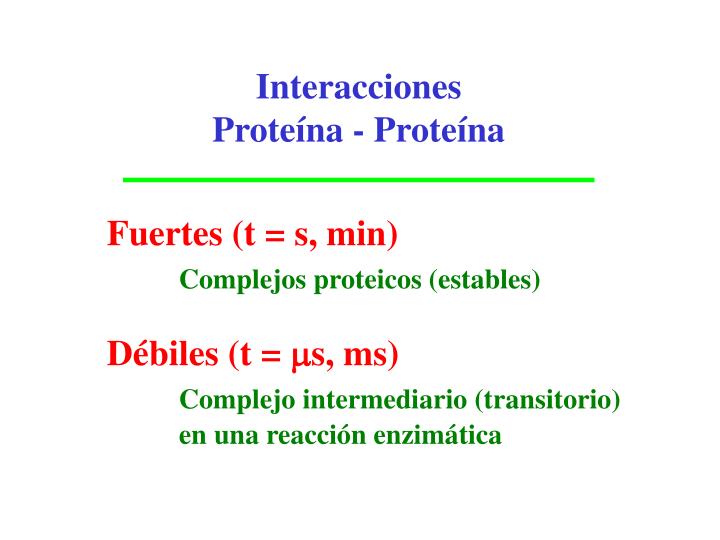
Interacciones Proteína - Proteína. Fuertes (t = s, min) Complejos proteicos (estables) Débiles (t = m s, ms) Complejo intermediario (transitorio) en una reacción enzimática. Interactions between functional group s. Schwikowski et al . (2000) Nature Biotec h. 18, 1257 - 1261.

E N D
Interacciones Proteína - Proteína Fuertes (t = s, min) Complejos proteicos (estables) Débiles (t = ms, ms) Complejo intermediario (transitorio) en una reacción enzimática
Interactions between functional groups Schwikowski et al.(2000) Nature Biotech. 18, 1257 - 1261
Interactions between proteins of different compartments Schwikowski et al.(2000) Nature Biotech. 18, 1257 - 1261
Yeast SH3 domains — which recognize proline-rich peptides — generated a network containing 394 interactions among 206 proteins Tong et al. (2002) Science 295, 321-324
An interaction map of the yeast proteome assembledfrom published interactions Schwikowski et al.(2000) Nature Biotech. 18, 1257 - 1261
Protein network inSaccharomyces cerevisiae ..\..\LINKS\Ho Nature(2002).pdf Ho et al. (2002) Nature 415, 180
Analysing protein interactions: Systematic identification of protein complexes in Saccharomyces cerevisiae by mass spectrometry Kumar & Snyder (2002) Nature 415, 123-124 Ho, Y et al. (2002) Nature 415, 180 - 183
How does a trimeric G protein on the inside of a cell membrane respond to activationby a transmembrane receptor? Trimeric (abg) G proteins relay signals from transmembrane receptors to intracellular enzymes and ion channels, thereby mediating vision, smell, taste and the actions of many hormones and neurotransmitters T. Iiri et al. (1998) Nature 394, 35-38
The GTPase cycle of trimeric G proteins The 'turn-on' step begins when the activatedreceptor (R*) associates with the trimer of (aGDPbg), causing dissociation of GDP. Then GTPbinds to the complex of R* with the trimer in its 'empty' state (aebg), and the resulting GTP-induced conformational change causes aGTP to dissociate from R* and from bg. After the 'turn-off' step (hydrolysis of bound GTP to GDP and inorganic phosphate, Pi), aGDP reassociateswithbg. T. Iiri et al. (1998) Nature 394, 35-38
Contacts between Gbg (left) and Ga-GDP (right) Red dashed linesindicate contacts that appear to be required for receptor activation but not for Ga–Gbg association; greendashed lines indicatecontacts that are important for both functions T. Iiri et al. (1998) Nature 394, 35-38
How does a trimeric G protein on the inside of a cell membrane respond to activationby a transmembrane receptor? Biomedical relevance: G-protein mutations in patients with hypertension andinherited endocrine disorders enhance or block signals from stimulated receptors. T. Iiri et al. (1998) Nature 394, 35-38
PARP-1: A Perpetrator of Apoptotic Cell Death Apoptotic cell death is triggered by activation of the nuclear enzyme poly(ADP-ribose) polymerase-1 (PARP-1). Through unknown mechanisms, PAR formation and NAD+ depletion may trigger a cascade of events. A. Chiarugi &M.A. Moskowitz (2002) Science 297, 200
light b6f Fd PS I Pc Cyt c6 PSI-driven Electron Transfer
Oxygen content of the earth's atmosphere 1 Berkner-Marshall Point (Terrestrial life) 0.1 Pasteur Point Atmospheric Level (fractions of 21% v/v) (O respiration) 2 0.01 EUKARYOTES Photosynthetic O production 2 0.001 3 1 4 2 0 9 Time (10 years ago) (Adapted from Peschek, 1996) PROKARYOTES
2- S SO 2- 4 Fe Availability Cu 3 1 4 2 0 9 Time (10 years ago) (Adapted from Williams & Silva, 1997)
Cu ligands: His-35 Cys-84 His-87 Met-92 Plastocyanin
Heme ligands: His-19 Met-61 Cytochrome c6
Organism Protein pI ___________________________________________________ Spinach Plastocyanin 4.2 Monoraphidium Plastocyanin 3.7 Cytochrome c6 3.6 Anabaena Plastocyanin 9.0 Cytochrome c6 9.0 Synechocystis Plastocyanin 5.5 Cytochrome c6 5.6 ____________________________________________________ Isoelectric point of cytochrome c6 and plastocyanin isolated from different organisms ___________________________________________________
Cytochrome c6 Plastocyanin De la Rosa et al. (2002) Bioelectrochemistry 55, 41-45
Photosynthetic organisms growing under controlled conditions
K K A R P r o t + P S I [ P r o t . . . P S I ] [ P r o t . . . P S I ] * r e d r e d r e d r e d r e d r e d 1 2 3 n n n h h h K ' K ' A R P r o t + P S I [ P r o t . . . P S I ] [ P r o t . . . P S I ] * r e d o x r e d o x r e d o x 1' 2' 3' k e t Routes n h a: 1 1' 2' 3' 4 n h P r o t + P S I 4 b: 1 2 2' 3' 4 o x r e d n h c: 1 2 3 3' 4 De la Rosa et al. (2002) Bioelectrochemistry 55, 41-45
KINETIC TYPES FOR THE REACTION MECHANISM Type I Protred + PSIox Protox + PSIred Type II Protred + PSIox[Protred ... PSIox] Protox + PSIred Type III Protred + PSIox[Protred ... PSIox] [Protred ... PSIox]* Protox + PSIred KINETIC TYPES FOR THE REACTION MECHANISM Type I Protred + PSIox Protox + PSIred Type II Protred + PSIox[Protred ... PSIox] Protox + PSIred Type III Protred + PSIox[Protred ... PSIox] [Protred ... PSIox]* Protox + PSIred KINETIC TYPES FOR THE REACTION MECHANISM Type I Protred + PSIox Protox + PSIred Type II Protred + PSIox[Protred ... PSIox] Protox + PSIred Type III Protred + PSIox[Protred ... PSIox] [Protred ... PSIox]* Protox + PSIred KINETIC TYPES FOR THE REACTION MECHANISM Type I Protred + PSIox Protox + PSIred Type II Protred + PSIox[Protred ... PSIox] Protox + PSIred Type III Protred + PSIox[Protred ... PSIox] [Protred ... PSIox]* Protox + PSIred Navarro et al. (1997) J. Biol. Inorg. Chem. 2, 11-22