Download
1 / 1
10 likes | 160 Views
3. 4. 3. 5. 1. 2. 5. Predictability and detectability of biogeographic changes in plant distributions Vincent P. Gutschick, Dept. of Biology, New Mexico State Univ., Las Cruces, NM 88003 and Global Change Consulting Consortium, Las Cruces, NM. B43B-1159.

E N D
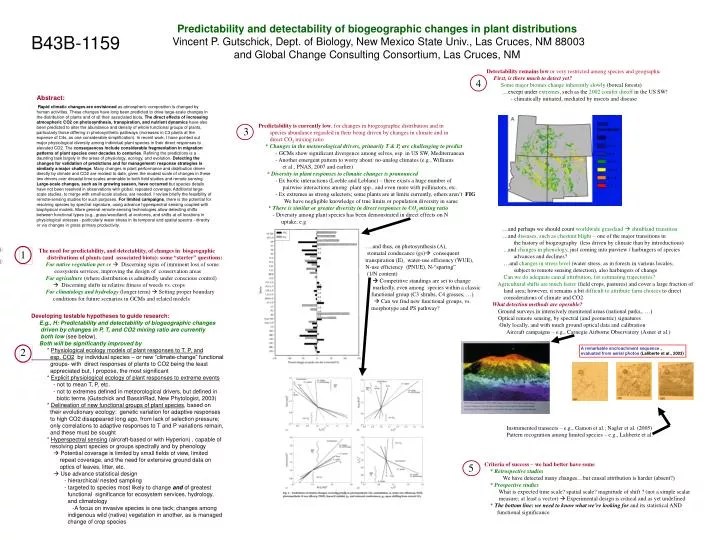
3 4 3 5 1 2 5 Predictability and detectability of biogeographic changes in plant distributions Vincent P. Gutschick, Dept. of Biology, New Mexico State Univ., Las Cruces, NM 88003 and Global Change Consulting Consortium, Las Cruces, NM B43B-1159 Detectability remains low or very restricted among species and geographic First, is there much to detect yet? Some major biomes change inherently slowly (boreal forests) …except under extremes, such as the 2002 conifer dieoff in the US SW! - climatically initiated, mediated by insects and disease …and perhaps we should count worldwide grassland shrubland transition …and diseases, such as chestnut blight – one of the major transitions in the history of biogeography (less driven by climate than by introductions) ...and changes in phenology, just coming into purview / harbingers of species advances and declines? …and changes in stress level (water stress, as in forests in various locales; subject to remote sensing detection), also harbingers of change Can we do adequate causal attribution, for estimating trajectories? Agricultural shifts are much faster (field crops, pastures) and cover a large fraction of land area; however, it remains a bit difficult to attribute farm choices to direct considerations of climate and CO2 What detection methods are operable? Ground surveys in intensively monitored areas (national parks,. …) Optical remote sensing, by spectral (and geometric) signatures Only locally, and with much ground optical data and calibration Aircraft campaigns – e.g., Carnegie Airborne Observatory (Asner et al.) Instrumented transects – e.g., Gamon et al.; Nagler et al. (2005) Pattern recognition among limited species – e.g., Laliberte et al. Abstract: Rapid climatic changes are envisioned as atmospheric composition is changed by human activities. These changes have long been predicted to drive large-scale changes in the distribution of plants and of all their associated biota. The direct effects of increasing atmospheric CO2 on photosynthesis, transpiration, and nutrient dynamics have also been predicted to alter the abundance and density of whole functional groups of plants, particularly those differing in photosynthetic pathways (increases in C3 plants at the expense of C4s, as one considerable simplification). In recent work, I have pointed out major physiological diversity among individual plant species in their direct responses to elevated CO2. The consequences include considerable fragmentation in migration patterns of plant species over decades to centuries. Refining the predictions is a daunting task largely in the areas of physiology, ecology, and evolution. Detecting the changes for validation of predictions and for management/ response strategies is similarly a major challenge. Many changes in plant performance and distribution driven directly by climate and CO2 are modest to date, given the modest scale of changes in these two drivers over decadal time scales amenable to both field studies and remote sensing. Large-scale changes, such as in growing season, have occurred but species details have not been resolved in observations with global, repeated coverage. Additional large-scale studies, to merge with small-scale studies, are needed. I review briefly the feasibility of remote-sensing studies for such purposes. For limited campaigns, there is the potential for resolving species by spectral signature, using advance hyperspectral sensing coupled with biophysical models. More general remote-sensing technologies allow detecting shifts between functional types (e.g., grass/woodland) at ecotones, and shifts at all locations in physiological stresses - particularly water stress in its temporal and spatial spectra - directly or via changes in gross primary productivity. Predictability is currently low, for changes in biogeographic distribution and in species abundance regarded in their being driven by changes in climate and in direct CO2 mixing ratio * Changes in the meteorological drivers, primarily T & P, are challenging to predict - GCMs show significant divergence among selves, esp. in US SW, Mediterranean - Another emergent pattern to worry about: no-analog climates (e.g., Williams et al., PNAS, 2007 and earlier) * Diversity in plant responses to climatic changes is pronounced - Ex biotic interactions (Loehle and Leblanc) – there exists a huge number of pairwise interactions among plant spp., and even more with pollinators, etc. - Ex extremes as strong selectors; some plants are at limits currently, others aren’t FIG We have negligible knowledge of true limits or population diversity in same * There is similar or greater diversity in direct responses to CO2 mixing ratio - Diversity among plant species has been demonstrated in direct effects on N uptake, e.g ….and thus, on photosynthesis (A), stomatal conducance (gs) consequent transpiration (E), water-use efficiency (WUE), N-use efficiency (PNUE), N-“sparing” (1/N content) Competitive standings are set to change markedly, even among species within a classic functional group (C3 shrubs, C4 grasses, …) Can we find new functional groups, vs. morphotype and PS pathway? The need for predictability, and detectablity, of changes in biogeogaphic distributions of plants (and associated biota): some “starter” questions: For native vegetation per se Discerning signs of imminent loss of some ecosystem services; improving the design of conservation areas For agriculture (where distribution is admittedly under conscious control) Discerning shifts in relative fitness of weeds vs. crops For climatology and hydrology (longer term) Setting proper boundary conditions for future scenarios in GCMs and related models Developing testable hypotheses to guide research: E.g., H: Predictability and detectability of biogeographic changes driven by changes in P, T, and CO2 mixing ratio are currently both low (see below), Both will be significantly improved by * Physiological ecology models of plant responses to T, P, and esp. CO2 by individual species – or new “climate-change” functional groups- with direct responses of plants to CO2 being the least appreciated but, I propose, the most significant * Explicit physiological ecology of plant responses to extreme events - not to mean T, P, etc. - not to extremes defined in meteorological drivers, but defined in biotic terms (Gutschick and BassiriRad, New Phytologist, 2003) * Delineation of new functional groups of plant species, based on their evolutionary ecology: genetic variation for adaptive responses to high CO2 disappeared long ago, from lack of selection pressure; only correlations to adaptive responses to T and P variations remain, and these must be sought * Hyperspectral sensing (aircraft-based or with Hyperion) , capable of resolving plant species or groups spectrally and by phenology Potential coverage is limited by small fields of view, limited repeat coverage, and the need for extensive ground data on optics of leaves, litter, etc. Use advance statistical design - hierarchical/ nested sampling - targeted to species most likely to change and of greatest functional significance for ecosystem services, hydrology, and climatology -A focus on invasive species is one tack; changes among indigenous wild (native) vegetation in another, as is managed change of crop species A remarkable encroachment sequence , evaluated from aerial photos (Laliberte et al., 2003) Criteria of success – we had better have some * Retrospective studies We have detected many changes…but causal attribution is harder (absent?) * Prospective studies What is expected time scale? spatial scale? magnitude of shift ? (not a simple scalar measure; at least a vector) Experimental design is critical and as yet undefined * The bottom line: we need to know what we’re looking for and its statistical AND functional significance




![Transient Mass-Loss Events in the PG 1159 [ WCE ] Central Star of Longmore 4](https://cdn1.slideserve.com/3355375/transient-mass-loss-events-in-the-pg-1159-wce-central-star-of-longmore-4-dt.jpg)
