Download

1 / 84
890 likes | 1.22k Views
Protein Structure and Function. CHAPTER3. Control of Protein Function. 3-0 Overview : Mechanisms of Regulation. Protein function in living cells is precisely regulated

E N D
3-0 Overview : Mechanisms of Regulation • Protein function in living cells is precisely regulated • Cells are densely packed (250,000 proteins/ a bacteria); regulation of protein function is essential to avoid chaos. (by localization,interaction with effector molecules and by the amount and lifetime of the active protein) • Proteins can be targeted to specific compartments and complexes - by signal sequences (specific amino-acid sequence), lipid tails and interaction domains When the protein is not in the location where it is needed, very often it is maintained in an inactive conformation.
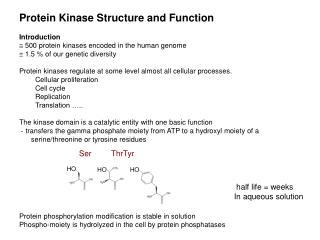
3-0 Overview : Mechanisms of Regulation • Protein activity can be regulated by binding of an effector and by covalent modification • Binding of effector induces conformational changes that produce inactive or active forms of the protein. • Post-translational covalent modification may either activate or inactivate the proteins. (phosphorylation, methyltation, acetylation, carbohydration, proteolytic cleavage…) • Signal amplification is an essential for the control of cell function and covalent modification is the way such amplification is usually achieved; kinase cascade, blood clotting
3-0 Overview : Mechanisms of Regulation • Protein activity may be regulated by protein quantity and lifetime • Amount of protein can be set by the level of transcription (promoter strength or transcription factor) • Amount of mRNA can be regulated by RNA degradation. • Amount of protein can by regulated by Ubiquitin-proteosome degradation.
3-0. Overview : Mechanisms of Regulation • A single protein may be subject to many regulatory influences • is achieved largely through signal transduction networks . • Ex)The cyclin-dependent protein kinases (CDK) to control the cell cycle are regulated by a number of different mechanisms.
3-0. Overview : Mechanisms of Regulation • Ckd and cyclin binding • → conformational change • → phosphorylation by CAK • → activation • further phosphorylation or dephosphotylation on other tyrosines finely regulate the activities Figure3-1. The cyclin-dependent protein kinases that control progression through the cell cycle are regulated by a number of different mechanisms
3-1. Protein Interaction Domains Different small domains → distinct binding specificities and funtions Phosphoserine and phosphothreonine motif Toll-like receptors - Interaction domains can be divided into distinct families whose members are related by sequence structure, and ligand binding properties. Figure3-2. Interaction domains
3-1. Protein Interaction Domains b-catenin Vesicle fusion Figure3-2. Interaction domains
3-1. Protein Interaction Domains IκBs mask NLS of NF-kB proteins; Figure3-2. Interaction domains
3-1. Protein Interaction Domains Figure3-2. Interaction domains
3-1. Protein Interaction Domains Figure3-2. Interaction domains
3-1. Protein Interaction Domains Figure3-2. Interaction domains
3-2. Regulation by Location • Protein function in the cell is context-dependent • Temporal and spatial control over a protein’s activity must be exercised. • Temporal control: regulating gene expression and protein lifetime. • Spatial control: location control. • Translocated by a specific organelle, a cargo vesicle. • Precise localization of protein is essential for proper protein function Various Kinases, Tem1 Figure3-3. The internal structure of cells
3-2. Regulation by Location There are several ways of targeting proteins in cells (a). By sequences in the protein itself (b-c). Covalent chemical modification of the protein (d). Binding to scaffold proteins Figure3-4. Mechanisms for targeting proteins
3-3. Control by pH and Redox Environment • - Protein function is modulated by the environment in which the protein operates • Changes in redox environment can greatly affect protein structure and function • : cysteine residues in proteins are usually fully reduced to –SH groups inside the cell but are readily oxidized to disulfide bond when secreted. • Changes in pH drastically alter protein structure and function • : Modulation of the surface charge of a protein by pH change could influence binding strength . • : charged group in their active sites can be changed by pH alteration (two Asp in active site). Active site open Figure3-5. Cathepsin D, endopeptidase conformational switching in endosome (pH ~5)
3-3. Control by pH and Redox Environment Proton pump Diphtheria toxin : B, T, A domain B domain : bind to a receptor in the target-cell membrane. A domain : kills cells by catalyzing the ADP-ribosylation of elongation factor 2 on the ribosome. T domain : deliver the catalytic domain into the cytoplasm of the target cell. Exposure to the reducing environment inside the endocytic vesicle breaks the disulfide bond between the A and B domains, releasing the toxic A domain. In low pH, the exposure of hydrophobic residues of the diphtheria toxin T domain results in its insertion into the endosomal membrane →As a channel, T domain transfers the A domain Figure3-6. Schematic representation of the mechanism by which diphtheria toxin kills a cell
Glutathione serves as a sulfhydryl buffer and an antioxidant
Structure of glutathione peroxidase 2GSH + RO-OH GSSG + H2O + ROH
3-4. Effector Ligands : Competitive Binding and Cooperativity Protein function can be controlled by effector ligands that bind competitively to ligand-binding or active sites - FEED BACK INHIBITION : the end product of the pathway acts as a competitive inhibitor of the first enzyme. Figure3-7. Competitive feedback inhibition
3-4. Effector Ligands : Competitive Binding and Cooperativity • Cooperative binding by effector ligands amplifies their effects • -Amplification: covalent modification, cooperativity • Cooperativity : positive cooperativity • negative cooperativity • Positive cooperativity : binding of one molecule of a ligand to a protein makes it easier for a second molecule of that ligand to bind. • Nagative cooperativity : binding of the second molecule is more difficult. Figure3-8. Cooperative ligand binding
3-5. Effector Ligands : Conformational Change and Allostery Effector molecules can cause conformational changes at distant sites • (a) The binding of successive effector molecules causes a sequential series of conformational changes form the initial state to the final state by induced fit. • subunits need not exist in the same conformation • substrate-binding causes increased substrate affinity in adjacent subunits • -conformational changes are not propagated to all subunits • (b). The effector can only bind to one of these forms, and its binding shifts the equilibrium in a concerted manner on favor of the bound form. • Allostery : from the Greek for “another structure” • Allostery activator • Allostery inhibitor Sequential model Concerted model Figure3-9. Two models of allosteric regulation
3-5. Effector Ligands : Conformational Change and Allostery • ATCase is an allosteric enzyme with regulatory and active sites on different subunits • Carbamoyl phosphate + Aspartate N-carbamoyl Aspartate, • ATCase is a hetero-oligomer. Six catalytic and six regulatory subunit. • Allosterically inhibited by cytidine triphosphate (end product, feedback inhibitor). • Allosterically activated by ATP(= activator). • Mutation of Y77F, the site away from active site stabilized the T state resulting in the inhibition of enzyme. CTP Figure3-10. Ligand-induced conformational change activates aspartate transcarbamoylase
3-5. Effector Ligands : Conformational Change and Allostery Binding of gene regulatory proteins to DNA is often controlled by ligand-induced conformational changes. - Co-activators and Co-repressors: small molecules, metal ions or proteins control binding of the activator or repressor to DNA. DtxR: specific repressor of Diphtheria toxin . Binding of DtxR to its operator sequence is controlled by the concentration of Fe2+ in the bacterial cell. Figure3-11. Iron binding regulates the repressor of the diphtheria toxin gene
3-6. Protein Switches Based on Nucleotide Hydrolysis Most protein switches are enzymes that catalyze the hydrolysis of a nucleoside triphosphate to the diphosphate - GTPase : major class of switch protein (G protein) - ATPase : usually associated with motor protein complexes or transporters ATPase GTPase -two-component response regulator: histidine kinase, response regulator proteins Why ATP or GTP are used for trigger of switch? Figure3-12. Structure of the core domains of a typical GTPase and an ATPase
3-6. Protein Switches Based on Nucleotide Hydrolysis • - Triphosphate-bound state = “on”, spring-loaded • Loss of gamma phosphate group → conformational change. • - Two hydrogen bonds in the each switch (Ⅰ and Ⅱ). a- and b- phosphates are bound to P-loop (GXXXXGKS/T) g-phospate is bound to both switch I and II (DXnT and DXXG respectively) Although common structural and functional features in switch proteins, many insertions of other domains in individual GTPases present various functions. Figure3-13. Schematic diagram of the universal switch mechanism of GTPases
3-7. GTPase Switches : Small Signaling G Proteins The switching cycle of nucleotide hydrolysis and exchange in G proteins is modulated by the binding of other proteins GTP hydrolysis rate is very low → GAP(GTPase-activating protein) increase the rate by 105 fold GDP release is conducted by GEF(guanidine-nucleotide exchange factors) Opening up the binding site Figure3-14. The switching cycle of the GTPase involves interactions with proteins that facilitate binding of GTP and stimulation of GTPase activity
Small GTPase Ras family: H-, N-,and K-ras, 21kDa, lipid attachment • Signal transduction by Ras is dependent on the GTP-bound state. A prolonged on state are found in up to 30% of human tumors. Reduction of GTP hydrolysis is caused by point mutations at 12, 13 or 61 resulting in uncontrolled cell growth and proliferation. Good target for anti-tumor therapy. • How the GAP facilitate GTP hydrolysis? - GAP insert an arginine side chain into the nucleotide-binding site of the GTPase. The positive charge on the side chain helps to stabilize the negative charge in the transition state for hydrolysis of the g-phosphate group of GTP
How the GEF facilitate GDP release? - GEF binding induces conformational changes in the P loop and switch regions of the GTPase while the rest of the structure is largely unchanged. The binding of the GEF sterically hinders the magnesium-binding site and interferes with the phosphate-binding region by insertion of an alpha helix into nucleotide binding site. When the GEF binds the GTPase, the phosphate groups are released first and the GEF is displaced upon binding of the entering GTP molecule. - After GDP has disassociated from the GTPase, GTP generally binds in its place, as the cytosolic ratio of GTP is much higher than GDP at 10:1. The binding of GTP to the GTPase results in the release of the GEF, which can then activate a new GTPase. Thus, GEFs both destabilize the GTPase interaction with GDP and stabilize the nucleotide free GTPase until a GTP molecule binds to it. From wikipedia
3-8. GTPase Switches : Signal Relay by Heterotrimeric GTPases • Heterotrimeric GTPase • α, β and γ subunit. • α subunit consist of the canonical G domain and an extra helical domain. • β and γ subunit are tightly associated with each other by coiled-coil interaction. • G protein associated with G protein coupled receptor(GPCR). • GDP-bound G protein bind to GPCR = “off” state. • When activated by ligand, these receptors act as GEF for their partner G protein. • When GDP is released and GTP binds, G protein dissociates from the GPCR. • In the absence of β and γ, α does not bind to GPCR.
Regulator of G-protein signaling proteins (RGS proteins) are responsible for the GTPase catalytic rate. How it increase the rate? • a subunit of G-protein has a “built-in” arginine residue in the extra helical domain that projects into the catalytic site. RGS proteins bind to the switch regions, reducing the flexibility and stabilization the transition state for hydrolysis. • Paticular RGS proteins regulate particular GPCRs; specificity • GPCRs are the most numerous receptors in all eukaryotic genome (1-5% of the total number of genes) • various ligands such as light, orants, lipids, peptide hormones. • 8 families
3-8. GTPase Switches : Signal Relay by Heterotrimeric GTPases GPCR = “Off” state WD40 Coiled-coil interaction Figure3-15. Hypothetical model of a heterotrimeric G protein in a complex with its G-protein-coupled receptor
3-9. GTPase Switches : Protein Synthesis EF-Tu(elongation factor) – GTP bound form Figure3-16. The switching cycle of the elongation factor EF-Tu delivers aminoacyl-tRNAs to the ribosome
3-10. Motor Protein Switches • Myosin is ATP-dependent nucleotide switches • ①ADP and Pi are bound to the heads as a result of hydrolysis of a bound ATP by the intrinsic ATPase activity of the catalytic core(blue). • ②Myosin head docking onto a specific binding site(green) on the actin thin filament(gray). • ③On actin docking Pi is released from the active site, and there is a conformational change in the head that causes the lever arm to swing to its “poststroke” ADP-bound position(red). • ④ADP dissociates and ATP binds to the active site and undergoes hydrolysis. ① ② ③ ④ Figure3-17. Models for the motor actions of muscle myosin and kinesin
3-10. Motor Protein Switches • Kinesin is ATP-dependent nucleotide switches • ①The “trailing” head has ADP bound and the “leading” head is empty and neither linker is docked tightly to the micro-tubule. • ②When ATP binds to the leading head, its linker adopts a conformation that as well as docking it firmly to the microtubule reverses its position and thus throws the trailing head forward by about 160Å towards the next binding site on the microtubule. • ③Binding also accelerates the release of ADP from this head, and during this time the ATP on the other head is hydrolyzed to ADP-Pi. • ④After ADP dissociates from the new leading head ATP binds in its turn, causing the linker to zipper onto the core. ① ② ③ ④ Figure3-17. Models for the motor actions of muscle myosin and kinesin
3-10. Motor Protein Switches Switch Ⅱ region of the motor protein kinesin Switch Ⅱ region of the motor protein Myosin Switch Ⅱ region of the G protein ATPase domains of motors and the GTPase domains of G proteins are different. Figure3-18. Structural and functional similarity between different families of molecular switches
The Inner Life of the Cell • http://www.youtube.com/watch?v=wJyUtbn0O5Y
3-12. Control of Protein Function by Phosphorylation • Protein function can by controlled by covalent modification • 50~90% of the proteins in the human body are post-tranlationally modified. • Phosphorylation, glycosylation, lipidation, and limited proteolysis…… • Most covalent modifications can change the location of the protein, or its activity, or its interatcions with other proteins and macromolecules. • Two phosphorylation effect : 1). Change the activity of the target protein 2). Provide a new recognition site for another protein to bind