Download
1 / 55
570 likes | 729 Views
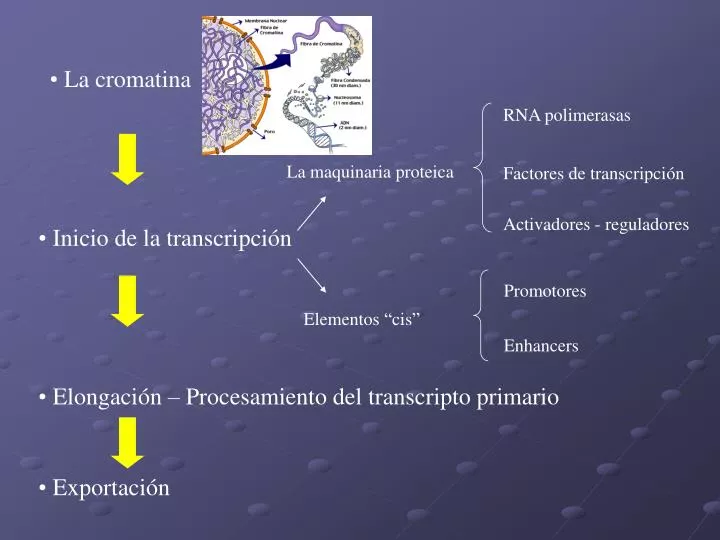
RNA polimerasas. Factores de transcripción. Activadores - reguladores. Promotores. Enhancers. La cromatina. La maquinaria proteica. Inicio de la transcripción. Elementos “ cis ”. Elongación – Procesamiento del transcripto primario. Exportación.

E N D
RNA polimerasas Factores de transcripción Activadores - reguladores Promotores Enhancers • La cromatina La maquinaria proteica • Inicio de la transcripción Elementos “cis” • Elongación – Procesamiento del transcripto primario • Exportación
A significant difference between the transcription of eukaryotic and prokaryotic mRNAs is that initiation at a eukaryotic promoter involves a large number of factors that bind to a variety of cis-acting elements.
The mechanism by which actinomycin D inhibits protein synthesis in animal cells HERBERT L. COOPER & RICHARD BRAVERMAN Section on Cellular and Molecular Physiology, Laboratory of Pathophysiology, National Cancer Institute, Bethesda, Maryland 20014 IN the course of studies on the regulation of protein synthesis during the activation of human lymphocytes by phytohaemagglutinin (PHA), it became necessary to determine whether an effect which occurred following treatment with actinomycin D (AMD) was secondary to the action of the drug on transcription, or resulted from a direct effect on translation. Using enucleated lymphocytes, we have shown that AMD has no effect on protein synthesis in the absence of the cell nucleus and conclude that AMD interferes with protein synthesis by its effect on transcription of RNA.
La transcripción de un gen eucariota codificante de proteínas es precedida por múltiples eventos • Decondensación del locus • Remodelado del nucleosoma • Modificaciones de histonas • Unión de activadores transcripcionales y coactivadores a enhancers y promotores • Posicionado de la maquinaria basal de transcripción al promotor
Las células eucarióticas poseen 3 tipos de RNA polimerasas en el núcleo Response to the bicyclic octapeptide α-amanitin. RNA polymerase II is rapidly inhibited by low concentrations of α-amanitin. RNA polymerase I is not inhibited. The response of RNA polymerase III to α-amanitin is less well conserved; in animal cells it is inhibited by high levels, but in yeast and insects it is not inhibited.
pol I rRNA ribosoma RNApol II DNA proteína mRNA pol III tRNA
La RNA pol II de levadura y de humano son virtualmente idénticas • 12 subunidades • 53 % de identidad • se pueden intercambiar subunidades (10)
Maquinaria de transcripción de la RNApol II DNA unwinding pol II RNA polymerization GTFs Promoter recognition Mediator Response to regulators
GTFs enables promoter recognition and initiation pol TATA 25 bp
General Transcription Factors TFII subunits masssimilarity to human B 1 37 52 % D (TBP) 1 21 81 % (TAFs) 10 662 E 2 109 53% F 2 159 51% H 9 570 62%
El complejo “mediator” es necesario para la regulación enhancer activator mediator pol TATA 25 bp Fue descubierta en levadura como una fracción cruda necesaria para la respuesta “in vitro”
Los factores de transcripción se suelen clasificar según el tipo de dominio de fijación al DNA que contienen Proteínas con “dedos de Zn” TFIIB, GAL4
Los factores de transcripción se suelen clasificar según el tipo de dominio de fijación al DNA que contienen Proteína con cremallera de leucina Proteína con hélice-bucle-hélice Proteínas de homeodominio c-fos and c-jun (the AP1 transcription factor), important regulators of normal development, as well as myc family members including myc, max, and mdx1. If they are overproduced or mutated in a vital area, they may generate cancer. These proteins interact with the DNA as dimers (homo- or hetero-) and are also called basic zipper proteins (bZips).
Ver simulación del proceso de transcripción en www.dnai.org
Incluye los elementos de ADN que se extienden 35 bp upstream y/o downstream del sitio de iniciación de la transcripción. Muchos interaccionan directamente con componentes de la maquinaria basal de transcripción. Núcleo del promotor (“core promoter”) Factores que son mínimamente esenciales para la transcripción (inclusive la RNA pol II) maquinaria basal de transcripción
Propiedades de los elementos promotores de la RNA polimerasa II (PROPERTIES OF RNA POLYMERASE II COREPROMOTER ELEMENTS)
TATA Box TATA Box(Goldberg-Hogness box) Secuencia TATAAA presente a 25 to 30 bp upstream de +1 • PREVALENCIA: En Drosophila 43% de 205 promotores
RECONOCIMIENTO DE TATA Roeder y colegas: transcription factor IID, TFIID es esencial para la transcripción a partir de promotores TATA. TFIID es un complejo TBP (proteína de reconocimiento) A binding site selection analysis identifiedthe sequence 5-TATATAAG-3 as the optimal TBP recognition sequence. TBP-associated factors (TAFs), actividades acetil transferasas quinasas, activadoras y conjugantes de ubiquitina
TATA +1 ATAT • TATA Box Rol de TATA en la direccionalidad de la transcripción +1 En un promotor que contiene sitios de unión distales para activadores La inversión del TATA box no produce un cambio en la dirección de la transcripción El principal determinante de la direccionalidad sería la disposición relativa de los sitios de unión de activadores, el TATA box y otros componentes del núcleo promotor TFIIB y RNA polymerasa II determinan la distancia a +1
Inr Initiator Element PyPy A(1) N T/A Py Py El elemento iniciador (Inr) fue definido como un elemento discreto del núcleo promotor que es funcionalmente similar a TATA box y puede actuar independientemente Espaciado TATA-INR y direccionalidad de la transcripción 15 or 20 bp se mantiene el sinergismo pero el +1 lo dicta el TATA box 25-30 bp los dos elementos actúan sinérgicamente > 30 bp actúan independientemente La inversión de Inr en el contexto de un promotor conteniendo sitios de unión para activadores, no cambia la dirección de transcripción
Inr • TFIIDinteracciona específicamente con Inr • La acción sinérgica de TATA y Inr correlaciona con la unión cooperativa de TFIID a los dos elementos • TFIIAes indispensable para la unión de TFIID a Inr • RNA polymerasa II reconoce ADN inespecíficamente, sin embargo posee una débil e intrínseca preferencia por secuencias de tipo Inr.
DPE Downstream Promoter Element (DPE) • El DPE actúa conjuntamente con Inr, • La secuencia principal de DPE está localizada a 28 to 32 ntdel A 1 en el motivo Inr • Un promotor dependiente de DPE contiene Inr y DPE • Mutaciones en cualquiera de los dos elementos resultan en la pérdida de unión de TFIIDy de actividad basal de transcripción
BRE TFIIB Recognition Element (BRE) TFIIB humano reconoce una secuencia de 7-bp: G/C G/C G/ACGCC El reconocimiento es mediado por un motivo HTH del C-terminal de TFIIB Este motivo no existe en levaduras ni plantas lo cual sugiere que BRE no contribuye a la regulación génica en ellos
CpG Islands CpG Islands El dinucleótido CpG es poco frecuente en genomas de vertebrados debido a que 5-metilcitosina puede transformarse en timina (por desaminación) el cual no es reparado • Existen segmentos de ADN de 0.5–2 kbp que poseen una alta densidad de dinucleótidos CpG. • El genoma humano contiene 29,000 islas CpG • En mamíferos la mitad de los promotores codificantes de proteínas están asociados con CpG • Las islas CpG carecen de TATA boxes, DPE o Inr . • Se caracterizan por la presencia de múltiples sitios de iniciación de la transcripción que abarcan una región de 100 bp o más • presentan múltiples sitios de unión para el factor de transcripción Sp1
These structures provide for the first time unequivocal evidence of the RNA polymerase II elongation complex • a discrete RNA exit groove located near the base of the largely unstructured carboxy-terminal domain (CTD) • accepts the nascent transcript as it is separated from the coding strand of the DNA template 10 nucleotides upstream of the transcript’s 3-end A maximum of 4 base pairs of unwound template DNA ahead of the 3-OH terminus of the growing transcript 9 base pair RNA-DNA hybrid that holds the newly synthesized transcript in register with the DNA template
The most notable structural difference between free and transcribing RNA polymerase II is the position of a large, 50 kDa hinged region referred to as the “clamp” In free polymeraseis in an open conformation that would allow easy entry of the DNA template into the catalytic site. In the elongating enzyme, closes tightly over the DNA and nascent transcript.
mRNA is synthesized in cells, on chromatin templates, at rates approaching 2000 nucleotides per minute. • Purified RNA polymerase II is capable of elongating RNA chains in vitro, even on naked DNA templates, at rates of only 100 to 300 nucleotides per minute
¿Cómo es el proceso de elongación? ¿Por qué se requieren de factores de elongación?
reinicio detención Pausa La RNA polimerasa II durante la elongación a cada paso de agregado de nucleótido oscila entre las conformaciones activa e inactiva (pausa) En algunos sitios del molde de ADN, la RNA pol “en pausa” es detenida, esa situación puede ser revertida sólo por factores de elongación (SII) La detención y la pausa sería el resultado de un movimiento en retroceso (2 – 4 nt) de la enzima y de la burbuja de transcripción El extremo 3´-OH no es aceptor del ribonucleótido trifosfato entrante. Luego de un tiempo la enzima espontáneamente se desliza hacia delante y el extremo se realinea con el ion Mg catalítico, continuando la síntesis. La RNA detenida resulta de un retroceso irreversible de 7 to 14 nucleótidos
TFIIF TFIIF was initially identified and purified as an essential RNA polymerase II initiation factor that is required for stable binding of polymerase to TFIID and TFIIB at the promoter and for entry of TFIIE and TFIIH into the preinitiation complex TFIIF and members of the ELL and Elongin families, can interact directly with elongating polymerase and increase its elongation rate by suppressing transient pausing by the enzyme at all or most steps of nucleotide addition Helping to maintain the 3-OH terminus of the nascent transcript in proper alignment with the polymerase catalytic site, thereby preventing backtracking by the enzyme.
CTD is essential for the functionof RNA polymerase II deletions that remove only a fraction of theheptapeptide repeats can be lethal phosphorylation of the CTD in elongation by RNApolymerase II • the CTDsof actively elongating polymerases are highly phosphorylated • polymerases containing hypophosphorylated CTDs preferentially enter thepreinitiation complex, where they are subsequently phosphorylated during orshortly after initiation
ELONGATOR AND FACT PROMOTE RNAPOLYMERASE II ELONGATION THROUGHCHROMATIN Elongator and FACT appear to promote RNA polymerase II elongation by modifying nucleosomes in ways that facilitate passage of polymerase through chromatin. Elongator is highly conserved from yeast to man. Notable among the Elongator subunits are Elp3, a histone acetyltransferase (HAT) of the GCN5-related N-acetyltransferase (GNAT) family, Mechanistic studies suggest that FACT stimulates RNA polymerase II elongation through nucleosomes by binding them and promoting removal of nucleosomal histones H2Aand H2B
THE RNA POLYMERASE II ELONGATION COMPLEX FUNCTIONS AS A PLATFORM THAT COORDINATES mRNA PROCESSING AND EXPORT
Formación del transcripto primario y su procesamiento durante la maduración del mRNA en células eucarióticas
Capping Capping of RNA polymerase II transcripts occurs when they reach a length of 25 to 30 nucleotides and requires the concerted action of three enzymes: an RNA triphosphatase, which removes the -phosphate at the 5-end of the nascent transcript; a guanylyltransferase, which transfers a GMP cap to the resulting diphosphate terminus of the nascent transcript; and a methyltransferase, which adds a methyl group to the N7 position of the GMP cap to form the transcript’s mature m7G(5)ppp(5)N cap structure. mammalian and yeast guanylyltransferase are recruited to transcribing RNA polymerase II through specific interactions with the phosphorylated CTD CTD phosphorylation by the TFIIH CTD kinase may provide a signal that links capping to the synthesis of nascent transcripts
Polyadenylation CPSF: cleavage and polyadenylation specificity factor CstF: cleavage stimulation factor the RNA polymerase CTD is required for efficient 3´ cleavage of transcripts prior to poly(A) addition by poly(A) polymerase
Los intrones pueden ser removidos por mecanismos diferentes Tipo I Tipo III Tipo II
splicing It is likely that the CTD recruits components of the splicing machinery to the RNA polymerase II elongation complex. SR and snRNP proteins have been shown to interact with phosphorylated RNA polymerase II and phosphorylated CTD peptides. The CTD could stimulate splicing at least in part by promoting assembly of early splicing intermediates, such as the U1 snRNP-5 splice site complex or the U2 snRNP-branch site complex
pol I rRNA ribosoma RNApol II DNA proteína mRNA pol III tRNA

