Download
1 / 1
10 likes | 179 Views
O.D. EFFECTS OF PARASITISM BY THE ECTOPARASITOID Bracon hebetor SAY (HYMENOPTERA: BRACONIDAE) ON HOST HEMOLYMPH PROTEINS OF THE MEDITERRANEAN FLOUR MOTH Ephestia kuehniella ZELLER (LEPIDOPTERA: PYRALIDAE). . Hülya Altuntaş 1 and A. Yavuz Kılıç 1.

E N D
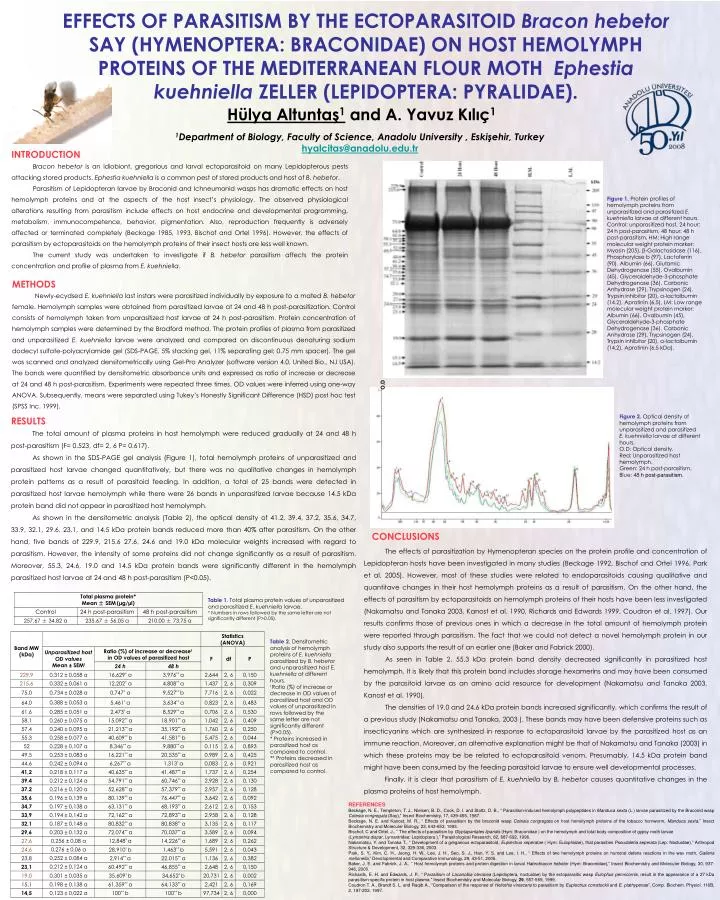
O.D EFFECTS OF PARASITISM BY THE ECTOPARASITOID Bracon hebetor SAY (HYMENOPTERA: BRACONIDAE) ON HOST HEMOLYMPH PROTEINS OF THE MEDITERRANEAN FLOUR MOTH Ephestia kuehniella ZELLER (LEPIDOPTERA: PYRALIDAE). Hülya Altuntaş1 and A. Yavuz Kılıç1 1Department of Biology, Faculty of Science, Anadolu University , Eskişehir, Turkey hyalcitas@anadolu.edu.tr INTRODUCTION Bracon hebetor is an idiobiont, gregarious and larval ectoparasitoid on many Lepidopterous pests attacking stored products. Ephestiakuehniella is a common pest of stored products and host of B. hebetor. Parasitism of Lepidopteran larvae by Braconid and Ichneumonid wasps has dramatic effects on host hemolymph proteins and at the aspects of the host insect’s physiology. The observed physiological alterations resulting from parasitism includeeffects on host endocrine and developmental programming, metabolism, immunocompetence, behavior, pigmentation. Also, reproduction frequently is adversely affected or terminated completely (Beckage 1985, 1993, Bischof and Ortel 1996). However, the effects of parasitism by ectoparasitoids on the hemolymph proteins of their insect hosts are less well known. The current study was undertaken to investigate if B. hebetor parasitism affects the protein concentration and profile of plasma from E. kuehniella. Figure 1. Protein profiles of hemolymph proteins from unparasitized and parasitized E. kuehniella larvae at different hours.Control: unparasitized host, 24 hour: 24 h post-parasitism, 48 hour: 48 h post-parasitism, HM: High range molecular weight protein marker: Myosin (205), β-Galactosidase (116), Phosphorylase b (97), Lactoferrin (90), Albumin (66), Glutamic Dehydrogenase (55), Ovalbumin (45), Glyceraldehyde-3-phosphate Dehydrogenase (36), Carbonic Anhydrase (29), Trypsinogen (24), Trypsin inhibitor (20), α-lactalbumin (14.2), Aprotinin (6.5), LM: Low range molecular weight protein marker: Albumin (66), Ovalbumin (45), Glyceraldehyde-3-phosphate Dehydrogenase (36), Carbonic Anhydrase (29), Trypsinogen (24), Trypsin inhibitor (20), α-lactalbumin (14.2), Aprotinin (6.5 kDa), METHODS Newly-ecydsed E. kuehniella last instars were parasitized individually by exposure to a mated B. hebetor female. Hemolymph samples were obtained from parasitized larvae at 24 and 48 h post-parasitization. Control consists of hemolymph taken fromunparasitized host larvae at 24 h post-parasitism. Protein concentration of hemolymph samples were determined by the Bradford method. The protein profiles of plasma from parasitized and unparasitized E. kuehniella larvae were analyzed and compared on discontinuous denaturing sodium dodecyl sulfate-polyacrylamide gel (SDS-PAGE, 5% stacking gel, 11% separating gel; 0.75 mm spacer). The gel was scanned and analyzed densitometrically using Gel-Pro Analyzer (software version 4.0, United Bio., NJ USA). The bands were quantified by densitometric absorbance units and expressed as ratio of increase or decrease at 24 and 48 h post-parasitism. Experiments were repeated three times. OD values were inferred using one-way ANOVA. Subsequently, means were separated using Tukey’s Honestly Significant Difference (HSD) post hoc test (SPSS Inc. 1999). Figure 2.Optical density of hemolymph proteins from unparasitized and parasitized E. kuehniellalarvae at different hours.O.D: Optical density.Red: Unparasitized host hemolymph.Green: 24 h post-parasitism.Blue: 48 h post-parasitism. RESULTS The total amount of plasma proteins in host hemolymph were reduced gradually at 24 and 48 hpost-parasitism (F= 0.523, df= 2,6 P= 0.617). As shown in the SDS-PAGE gel analysis (Figure 1), total hemolymph proteins of unparasitized and parasitized host larvae changed quantitatively, but there was no qualitative changes in hemolymph protein patterns as a result of parasitoid feeding. In addition, a total of 25 bands were detected in parasitized host larvae hemolymph while there were 26 bands in unparasitized larvae because 14.5 kDa protein band did not appear in parasitized host hemolymph. As shown in the densitometric analysis (Table 2), the optical density of 41.2, 39.4, 37.2, 35.6, 34.7, 33.9, 32.1, 29.6, 23.1, and 14.5 kDa protein bands reduced more than 40% after parasitism. On the other hand, five bands of 229.9, 215.6 27.6, 24.6 and 19.0 kDa molecular weights increased with regard to parasitism. However,the intensity of some proteins did not change significantly as a result of parasitism. Moreover, 55.3, 24.6, 19.0 and 14.5 kDa protein bands were significantly different in the hemolymph parasitized host larvae at 24 and 48 h post-parasitism (P<0.05). CONCLUSIONS The effects of parasitization by Hymenopteran species on the protein profile and concentration of Lepidopteran hosts have been investigated in many studies (Beckage 1992, Bischof and Ortel 1996, Park et al. 2005). However, most of these studies were related to endoparasitoids causing qualitative and quantitave changes in their host hemolymph proteins as a result of parasitism. On the other hand, the effects of parasitism by ectoparasitoids on hemolymph proteins of their hosts have been less investigated (Nakamatsu and Tanaka 2003, Kanost et al. 1990, Richards and Edwards 1999, Coudron et al. 1997). Our results confirms those of previous ones in which a decrease in the total amount of hemolymph protein were reported through parasitism. The fact that we could not detect a novel hemolymph protein in our study also supports the result of an earlier one (Baker and Fabrick 2000). As seen in Table 2, 55.3 kDa protein band density decreased significantly in parasitized host hemolymph. It is likely that this protein band includes storage hexamerins and may have been consumed by the parasitoid larvae as an amino acid resource for development (Nakamatsu and Tanaka 2003, Kanost et al. 1990). The densities of 19.0 and 24.6 kDa protein bands increased significantly, which confirms the result of a previous study (Nakamatsu and Tanaka, 2003 ). These bands may have been defensive proteins such as insecticyanins which are synthesized in response to ectoparasitoid larvae by the parasitized host as an immune reaction. Moreover, an alternative explanation might be that of Nakamatsu and Tanaka (2003) in which these proteins may be be related to ectoparasitoid venom. Presumably, 14.5 kDa protein band might have been consumed by the feeding parasitoid larvae to ensure well developmental processes. Finally, it is clear that parasitism of E. kuehniella by B. hebetor causes quantitative changes in the plasma proteins of host hemolymph. Table 1. Total plasma protein values of unparasitized and parasitized E. kuehniella larvae. * Numbers in rows followed by the same letter are not significantly different (P>0.05). Table 2. Densitometric analysis of hemolymph proteins of E. kuehniella parasitized by B. hebetor and unparasitized host E. kuehniella at different hours. IRatio (%) of increase or decrease in OD values of parasitized host and OD values of unparasitized in rows followed by the same letter are not significantly different (P>0.05). * Proteins increased in parasitized host as compared to control. ** Proteins decreased in parasitized host as compared to control. REFERENCES Beckage, N. E., Templeton, T. J., Nielsen, B. D., Cook, D. I. and Stoltz, D. B., “ Parasitism-induced hemolymph polypeptides in Manduca sexta (L.) larvae parasitized by the Braconid wasp Cotesia congregata (Say),” Insect Biochemistry, 17, 439-455, 1987. Beckage, N. E. and Kanost, M. R., “ Effects of parasitism by the braconid wasp Cotesia congregata on host hemolymph proteins of the tobacco hornworm, Manduca sexta,” Insect Biochemistry and Moleculer Biology, 23, 643-653, 1993. Bischof, C and Ortel, J., “ The effects of parasitism by Glyptapantales liparidis (Hym: Braconidae ) on the hemolymph and total body composition of gypsy moth larvae (Lymantriadispar, Lymantridae: Lepidoptera ),” Parasitological Research, 82, 687-692, 1996. Nakamatsu, Y. and Tanaka T., “ Development of a gregarious ectoparasitoid, Euplectrus seperatae ( Hym: Eulophidae), that parasities Pseudaletia seperata (Lep: Noctuidae),” Arthropod Structure & Development,32, 329-336, 2003. Park, S. Y., Kim, C. H., Jeong, H. W., Lee, J. H., Seo, S. J., Han, Y. S. and Lee, I. H., “ Effects of two hemolymph proteins on humoral defens reactions in the wax moth, Galleria mellonella,” Devolopmental and Comparative Immunology, 29, 43-51, 2005. Baker, J. E. and Fabrick, J. A., “ Host hemolymph proteins and protein digestion in larval Habrobracon hebetor (Hym: Braconidae),” Insect Biochemistry and Moleculer Biology, 30, 937-946, 2000. Richards, E. H. and Edwards, J. P., “ Parasitism of Lacanobia oleracea (Lepidoptera, noctuidae) by the ectoparasitic wasp Eulophus pennicornis, result in the appearance of a 27 kDa parasitism-specific protein in host plasma,” Insect Biochemistry and Molecular Biology, 29, 557-569, 1999. Coudron T. A., Brandt S. L. and Raqib A., “Comparison of the response of Heliothis virescens to parasitism by Euplectrus comstockii and E. plathypenae”, Comp. Biochem. Physiol. 116B, 2, 197-202, 1997.


