Download
1 / 1
10 likes | 135 Views
EVIDENCE FOR THE INVOLVEMENT OF RYANODINE RECEPTORS IN Ca 2+ RELEASE BY GROUP I METABOTROPIC GLUTAMATE RECEPTORS IN CULTURED RAT HIPPOCAMPAL NEURONS Aidan Kaar, John Leahy, Keith O'Donoghue & Mark G. Rae. Department of Physiology, University College Cork, Ireland. INTRODUCTION. 1.43.

E N D
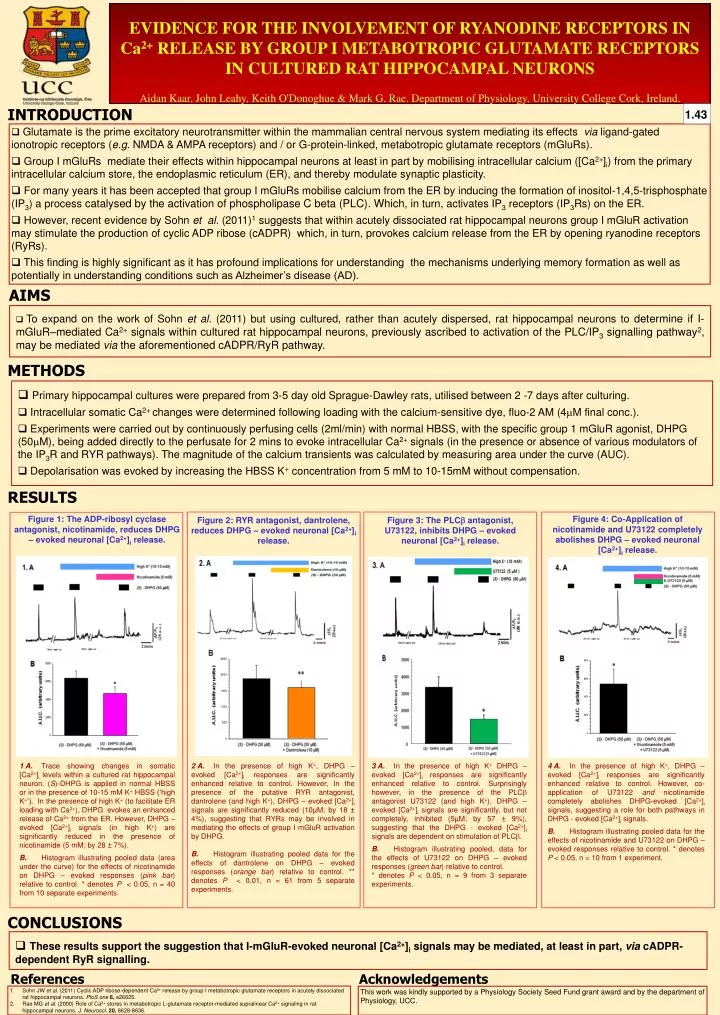
EVIDENCE FOR THE INVOLVEMENT OF RYANODINE RECEPTORS IN Ca2+ RELEASE BY GROUP I METABOTROPIC GLUTAMATE RECEPTORS IN CULTURED RAT HIPPOCAMPAL NEURONS Aidan Kaar, John Leahy, Keith O'Donoghue & Mark G. Rae. Department of Physiology, University College Cork, Ireland. INTRODUCTION 1.43 • Glutamate is the prime excitatory neurotransmitter within the mammalian central nervous system mediating its effects via ligand-gated ionotropic receptors (e.g. NMDA & AMPA receptors) and / or G-protein-linked, metabotropic glutamate receptors (mGluRs). • Group I mGluRs mediate their effects within hippocampal neurons at least in part by mobilising intracellular calcium ([Ca2+]i) from the primary intracellular calcium store, the endoplasmic reticulum (ER), and thereby modulate synaptic plasticity. • For many years it has been accepted that group I mGluRs mobilise calcium from the ER by inducing the formation of inositol-1,4,5-trisphosphate (IP3) a process catalysed by the activation of phospholipase C beta (PLC). Which, in turn, activates IP3 receptors (IP3Rs) on the ER. • However, recent evidence by Sohn et al. (2011)1 suggests that within acutely dissociated rat hippocampal neurons group I mGluR activation may stimulate the production of cyclic ADP ribose (cADPR) which, in turn, provokes calcium release from the ER by opening ryanodine receptors (RyRs). • This finding is highly significant as it has profound implications for understanding the mechanisms underlying memory formation as well as potentially in understanding conditions such as Alzheimer’s disease (AD). AIMS • To expand on the work of Sohn et al. (2011) but using cultured, rather than acutely dispersed, rat hippocampal neurons to determine if I-mGluR–mediated Ca2+ signals within cultured rat hippocampal neurons, previously ascribed to activation of the PLC/IP3 signalling pathway2, may be mediated via the aforementioned cADPR/RyR pathway. METHODS • Primary hippocampal cultures were prepared from 3-5 day old Sprague-Dawley rats, utilised between 2 -7 days after culturing. • Intracellular somatic Ca2+ changes were determined following loading with the calcium-sensitive dye, fluo-2 AM (4mM final conc.). • Experiments were carried out by continuously perfusing cells (2ml/min) with normal HBSS, with the specific group 1 mGluR agonist, DHPG (50mM), being added directly to the perfusate for 2 mins to evoke intracellular Ca2+ signals (in the presence or absence of various modulators of the IP3R and RYR pathways). The magnitude of the calcium transients was calculated by measuring area under the curve (AUC). • Depolarisation was evoked by increasing the HBSS K+ concentration from 5 mM to 10-15mM without compensation. RESULTS Figure 1: The ADP-ribosyl cyclase antagonist, nicotinamide, reduces DHPG – evoked neuronal [Ca2+]i release. Figure 4: Co-Application of nicotinamide and U73122 completely abolishes DHPG – evoked neuronal [Ca2+]i release. Figure 2: RYR antagonist, dantrolene, reduces DHPG – evoked neuronal [Ca2+]i release. Figure 3: The PLCb antagonist, U73122, inhibits DHPG – evoked neuronal [Ca2+]i release. 1 A. Trace showing changes in somatic [Ca2+]i levels within a cultured rat hippocampal neuron. (S)-DHPG is applied in normal HBSS or in the presence of 10-15 mM K+ HBSS (‘high K+’). In the presence of high K+ (to facilitate ER loading with Ca2+), DHPG evokes an enhanced release of Ca2+ from the ER. However, DHPG – evoked [Ca2+]i signals (in high K+) are significantly reduced in the presence of nicotinamide (5 mM; by 28 ± 7%). B. Histogram illustrating pooled data (area under the curve) for the effects of nicotinamide on DHPG – evoked responses (pink bar) relative to control. * denotes P < 0.05, n = 40 from 10 separate experiments. 2 A. In the presence of high K+, DHPG – evoked [Ca2+]i responses are significantly enhanced relative to control. However, In the presence of the putative RYR antagonist, dantrolene (and high K+), DHPG – evoked [Ca2+]i signals are significantly reduced (10µM; by 18 ± 4%), suggesting that RYRs may be involved in mediating the effects of group I mGluR activation by DHPG. B. Histogram illustrating pooled data for the effects of dantrolene on DHPG – evoked responses (orange bar) relative to control. ** denotes P < 0.01, n = 61 from 5 separate experiments. 3 A. In the presence of high K+ DHPG – evoked [Ca2+]i responses are significantly enhanced relative to control. Surprisingly however, in the presence of the PLCb antagonist U73122 (and high K+), DHPG – evoked [Ca2+]i signals are significantly, but not completely, inhibited (5µM; by 57± 9%), suggesting that the DHPG - evoked [Ca2+]i signals are dependent on stimulation of PLCb. B. Histogram illustrating pooled, data for the effects of U73122 on DHPG – evoked responses (green bar) relative to control. * denotes P < 0.05, n = 9 from 3 separate experiments. 4 A. In the presence of high K+, DHPG – evoked [Ca2+]i responses are significantly enhanced relative to control. However, co-application of U73122 andnicotinamide completely abolishes DHPG-evoked [Ca2+]isignals, suggesting a role for both pathways in DHPG - evoked [Ca2+]isignals. B. Histogram illustrating pooled data for the effects of nicotinamide and U73122 on DHPG – evoked responses relative to control. * denotes P < 0.05, n = 10 from 1 experiment. CONCLUSIONS • These results support the suggestion that I-mGluR-evoked neuronal [Ca2+]i signals may be mediated, at least in part, via cADPR-dependent RyR signalling. References Acknowledgements 1. Sohn JW et al. (2011) Cyclic ADP ribose-dependent Ca2+ release by group I metabotropic glutamate receptors in acutely dissociated rat hippocampal neurons. PloS one6, e26625. 2. Rae MG et al. (2000) Role of Ca2+ stores in metabotropic L-glutamate receptor-mediated supralinear Ca2+ signaling in rat hippocampal neurons. J. Neurosci.20, 8628-8636. This work was kindly supported by a Physiology Society Seed Fund grant award and by the department of Physiology, UCC.






