Download
1 / 1
10 likes | 119 Views
a. IP. b. DIN. b. a. a. a, b. pmoles/million cells. c. b. CLA + IP. CLA + IP. IP. IP. a. control. X-ALD. a. b. b. CD 16:2 from 10,12. a. a. b. b. CD 16:2 from 9,11. nmoles/10 million cells. CLA. CLA. CLA+ IP. CLA + IP. control. X-ALD.

E N D
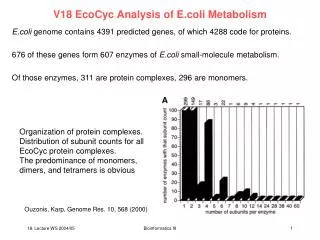
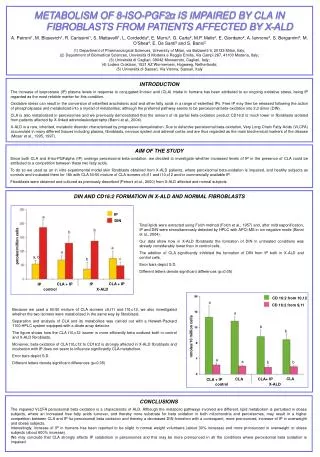
a IP b DIN b a a a, b pmoles/million cells c b CLA + IP CLA + IP IP IP a control X-ALD a b b CD 16:2 from 10,12 a a b b CD 16:2 from 9,11 nmoles/10 million cells CLA CLA CLA+ IP CLA + IP control X-ALD METABOLISM OF 8-ISO-PGF2α IS IMPAIRED BY CLA IN FIBROBLASTS FROM PATIENTS AFFECTED BY X-ALD A. Petroni1, M. Blasevich1, R. Carissimi1, S. Mattavelli1, L. Cordeddu2, E. Murru2, G. Carta2, M.P. Melis2, E. Giordano2, A. Iannone3, S. Bergamini3, M. O’Shea4, E. De Santi5 and S. Banni2 (1) Department of Pharmacological Sciences, University of Milan, via Balzaretti 9, 20133 Milan, Italy; (2) Department of Biomedical Sciences, Università di Modena e Reggio Emilia, Via Campi 287, 41100 Modena, Italy; (3) Università di Cagliari, 09042 Monserrato, Cagliari, Italy; (4) Loders Croklaan, 1521 AZ Wormerveer, Hogeweg, Netherlands; (5) Università di Sassari, Via Vienna, Sassari, Italy INTRODUCTION The increase of isoprostane (IP) plasma levels in response to conjugated linoleic acid (CLA) intake in humans has been attributed to an ongoing oxidative stress, being IP regarded as the most reliable marker for this condition. Oxidative stress can result in the conversion of esterified arachidonic acid and other fatty acids in a range of esterified IPs. Free IP may then be released following the action of phospholipases and metabolized into a myriad of metabolites, although the preferred pathway seems to be peroxisomal beta-oxidation into 2,3 diinor (DIN). CLA is also metabolized in peroxisomes and we previously demonstrated that the amount of its partial beta-oxidation product CD16:2 is much lower in fibroblasts isolated from patients affected by X-linked adrenoleukodystrophy (Banni et al., 2004). X-ALD is a rare, inherited, metabolic disorder characterised by progressive demyelination. Due to defective peroxisomal beta-oxidation, Very Long Chain Fatty Acids (VLCFA) accumulate in many different tissues including plasma, fibroblasts, nervous system and adrenal cortex and are thus regarded as the main biochemical markers of the disease (Moser et al., 1995, 1997). AIM OF THE STUDY Since both CLA and 8-iso-PGFalpha (IP) undergo peroxisomal beta-oxidation, we decided to investigate whether increased levels of IP in the presence of CLA could be attributed to a competition between these two fatty acids. To do so we used as an in vitro experimental model skin fibroblasts obtained from X-ALD patients, where peroxisomal beta-oxidation is impaired, and healthy subjects as controls and incubated them for 18h with CLA 50:50 mixture of CLA isomers c9,t11 and t10,c12 and/or commercially available IP. Fibroblasts were obtained and cultured as previously described (Petroni et al., 2000) from X-ALD affected and normal subjects. DIN AND CD16:2 FORMATION IN X-ALD AND NORMAL FIBROBLASTS Total lipids were extracted using Folch method (Folch et al., 1957) and, after mild saponification, IP and DIN were simoultaneously detected by HPLC with APCI-MS in ion negative mode (Banni et al., 2004). Our data show how in X-ALD fibroblasts the formation of DIN in untreated conditions was already considerably lower than in control cells. The addition of CLA significantly inhibited the formation of DIN from IP both in X-ALD and control cells. Error bars depict S.D. Different letters denote significant differences (p<0.05) Because we used a 50:50 mixture of CLA isomers c9,t11 and t10,c12, we also investigated whether the two isomers were metabolized in the same way by fibroblasts. Separation and analysis of CLA and its metabolites was carried out with a Hewlett-Packard 1100 HPLC system equipped with a diode array detector. The figure shows how the CLA t10,c12 isomer is more efficiently beta oxidized both in control and X-ALD fibroblasts. Moreover, beta-oxidation of CLA t10,c12 to CD16:2 is strongly affected in X-ALD fibroblasts and incubation with IP does not seem to influence significantly CLA metabolism. Error bars depict S.D. Different letters denote significant differences (p<0.05) CONCLUSIONS The impaired VLCFA peroxisomal beta oxidation is a characteristic of ALD. Although the metabolic pathways involved are different, lipid metabolism is perturbed in obese subjects, where an increased free fatty acids turnover, and thereby more substrate for beta oxidation in both mitochondria and peroxisomes, may result in a higher competition between CLA and IP for peroxisomal beta oxidation and thereby a decreased DIN formation with a consequent, more pronounced, increase of IP in overweight and obese subjects. Interestingly, increase of IP in humans has been reported to be slight in normal weight volunteers (about 30% increase) and more pronounced in overweight or obese subjects (about 600% increase). We may conclude that CLA strongly affects IP catabolism in peroxisomes and this may be more pronounced in all the conditions where peroxisomal beta oxidation is impaired.