Download

1 / 14
150 likes | 474 Views
CITRIC ACID CYCLE -Anaplerosis. Reading: Harper’s Biochemistry Chapter 18 Lehninger Principles of Biochemistry 3rd Ed. pp. 584-592. OBJECTIVES. To understand the citric acid cycle as both a source and “sink” for carbon compounds involved in other metabolic pathways.

E N D
CITRIC ACID CYCLE-Anaplerosis Reading: • Harper’s Biochemistry Chapter 18 • Lehninger Principles of Biochemistry 3rd Ed. pp. 584-592
OBJECTIVES • To understand the citric acid cycle as both a source and “sink” for carbon compounds involved in other metabolic pathways. • To understand anaplerosis as a mechanism for regulating the flow of intermediates in the citric acid cycle. • To understand how the citric acid cycle is essential for supplying acetyl-CoA to the cytoplasm for use in fatty acid biosynthesis.
In aerobic organisms, the citric acid cycle is an amphibolic pathway, one that serves in both catabolic and anabolic processes. • Besides its role in the oxidative catabolism of carbohydrates, fatty acids, and amino acids, the cycle provides precursors for many biosynthetic pathways, through reactions that served the same purpose in anaerobic ancestors. • Ketoglutarate and oxaloacetate can serves as precursors for glutamate and aspartate, respectively, by simple transamination, which themselves can act as precursors for other amino acids and nucleotides. • Oxaloacetate can be converted to glucose in gluconeogenesis. • Succinyl-CoA is a central intermediate of heme groups.
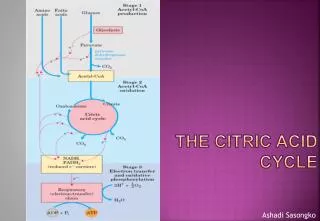
Role of the citric acid cycle in anabolism. Intermediates of the citric acid cycle are drawn off as precursors in many biosynthetic pathways. Shown in red are four anaplerotic reactions that replenish depleted cycle intermediates
Anaplerotic reactions replenish intermediates in citric acid cycle • As intermediates are removed to serve as biosynthetic precursors, they are replenished by anaplerotic reactions. • Under normal circumstances, removal and replenishment are in dynamic balance so intermediates stay almost constant.
Most significant is the formation of oxaloacetate by pyruvate carboxylase ATP + CO2 + H2O + pyruvateoxaloacetate + ADP + Pi • Pyruvate carboxylase is a regulatory enzyme and is virtually inactive in the absence of acetyl-CoA, its positive allosteric activator. Whenever acetyl-CoA, the fuel for the citric acid cycle, is in excess, it stimulates the pyruvate carboxylase reaction to make more oxaloacetate, enabling the cycle to proceed. • Pyruvate carboxylase has biotin as a prosthetic group. It is a specialized carrier of one carbon groups in their most oxidized form, CO2. • Biotin is required in the human diet, it is abundant in many foods and made by intestinal bacteria. Biotin deficiency is rare but can happen when large quantities of raw eggs are consumed as avidin in egg white is a tight biotin binder.
Regulation of acetyl-CoA production by the pyruvate dehydrogenase complex • The pyruvate dehydrogenase complex of vertebrates is strongly inhibited by ATP, by acetyl-CoA, and by NADH, the products of the reaction catalyzed by the complex. • When long chain fatty acids are available, and can provide acetyl-CoA via B-oxidation, pyruvate oxidation is inhibited. • When too little acetate flows through the cycle, AMP, CoA, and NAD+ all accumulate and allosterically activate the pyruvate dehydrogenase complex. • Pyruvate dehydrogenase is inhibited by reversible serine phosphorylation - the kinase responsible is allosterically activated by ATP.
Regulation of the citric acid cycle at its three exergonic steps • The flow of metabolites through the citric acid cycle is under stringent regulation. • Three major factors govern the rate of flux: substrate availability, inhibition by accumulating products, and allosteric feedback inhibition of the enzymes that catalyze early steps in the cycle. • Each of the three strongly exergonic steps - those catalyzed by citrate synthesis, isocitrate dehydrogenase, and -ketoglutarate dehydrogenase can become the rate-limiting step under specific conditions: -citrate synthase can limit the rate of citrate formation if substrates (acetyl-CoA and oxaloacetate) are at low level -high NADH/NAD+ ratio inhibits both dehydrogenase reactions -product accumulation inhibits all three steps -In muscle Ca2+, the signal for contraction and increased energy demand, activates isocitrate, -ketoglutarate, and pyruvate dehydrogenases
The citric acid cycle takes part in fatty acid biosynthesis • Acetyl-CoA is a major building block for long-chain fatty acid synthesis (in non-ruminants; in ruminants, acetyl-CoA is derived from acetate). • Since pyruvate dehydrogenase is a mitochondrial enzyme and the enzymes needed for fatty acid biosynthesis are extramitrochondrial, the acetyl-CoA is recovered as citrate, cleaved back to acetyl-CoA in the cytosol by ATP-citrate lyase
The glycoxylate cycle in plants • Vertebrates cannot convert fatty acids or acetate to carbohydrates • In many organisms other than vertebrates, the glyoxylate cycle serves as a mechanism for converting acetate to carbohydrate
Summary • The citric acid cycle is the final pathway for the oxidation of carbohydrate, lipid, and protein. It catalyzes the combination of their common metabolite, acetyl-CoA, with oxaloacetate to form citrate. Through a series of dehydrogenations and decarboxylations, citrate is degraded, producing reducing equivalents in the form of NADH and FADH2, releasing CO2, and regenerating oxaloacetate. • The overall rate of the citric acid cycle is controlled by the rate of conversion of pyruvate to acetyl-CoA and by the flux through citrate synthase, isocitrate dehydrogenase, and -ketoglutarate dehydrogenase. These fluxes are largely determined by the concentrations of substrates and products; the end products ATP and NADH are inhibitory.
Summary • Citric acid cycle intermediates are also used as precursors in the biosynthesis of amino acids and other biomolecules. These intermediates are replenished by anaplerotic reactions catalyzed by pyruvate decarboxylase, PEP carboxykinase, PEP carboxylase, and malic enzyme • The glyoxylate cycle in plants and some microorganisms bypasses the two decarboxylation steps of the citric acid cycle and makes possible the net formation of succinate and oxaloacetate from acetyl-CoA, glucose formation from fatty acids or acetate.














![Tricarboxylic acid cycle (TCA Cycle) [Kreb’s cycle] [Citric acid cycle]](https://cdn3.slideserve.com/6696193/tricarboxylic-acid-cycle-tca-cycle-kreb-s-cycle-citric-acid-cycle-dt.jpg)



