Download
1 / 33
360 likes | 1.06k Views
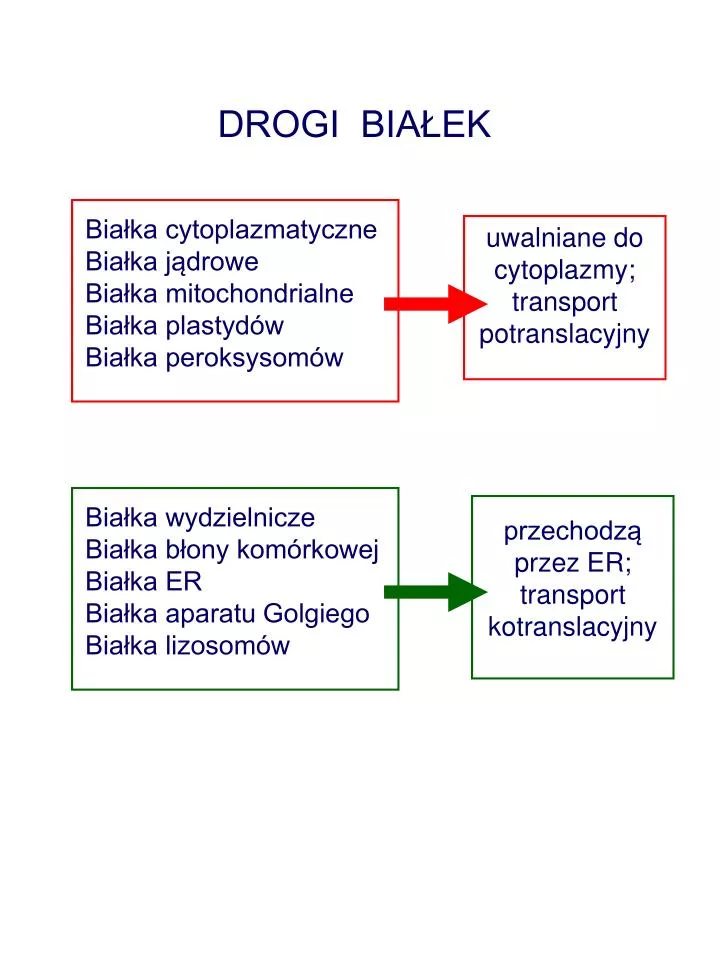
DROGI BIAŁEK. Białka cytoplazmatyczne Białka jądrowe Białka mitochondrialne Białka plastydów Białka peroksysomów. uwalniane do cytoplazmy; transport potranslacyjny. Białka wydzielnicze Białka błony komórkowej Białka ER Białka aparatu Golgiego Białka lizosomów. przechodzą przez ER;

E N D
DROGI BIAŁEK Białka cytoplazmatyczne Białka jądrowe Białka mitochondrialne Białka plastydów Białka peroksysomów uwalniane do cytoplazmy; transport potranslacyjny Białka wydzielnicze Białka błony komórkowej Białka ER Białka aparatu Golgiego Białka lizosomów przechodzą przez ER; transport kotranslacyjny
Sekwencja sygnałowa specyficzna dla białek mitochondrialnch
DROGI BIAŁEK mRNA + rybosomy polisomy wolne białka I grupy związane z ER białka II grupy http://wing-keung.tripod.com/cellbio2/cellbio2e.jpg
George Palade – 1974 - nagroda Nobla wraz z Albertem Claude i Christianem de Duve za odkrycia dotyczące strukturalnej i funkcjonalnej organizacji komórki. • George Palade (m.in.) lata 50. i 60. • odkrycie rybosomów (ciałka Palade’a; ang. Palade granules) • badania nad mechanizmem wydzielaniazymogenów przez komórki trzustki. Pytanie: Czy istnieje jedna czy dwie klasy rybosomów? Czy wszystkie rybosomy zdolne są do wiązania się z ER, czy też może rybosomy występujące w cytozolu są inne niż te związane z ER?
RER versus SER Czy szorstkie retikulum różni się od gładkiego? Czy szorstkie retikulum to jest po prostu gładkie retikulum z przyłączonymi rybosomami, czy to inna struktura, w jakiś jeszcze inny sposób różniąca się od gładkiego ER? INNA Wyizolowano szereg białek (co najmniej 20), które występują w szorstkim, a nie występują w gładkim ER. Część z tych białek jest zaangażowana w transport i potranslacyjne modyfikacje białek w obrębie ER.
Skąd wiadomo, że białkawydzielnicze wędrują do retikulum? Badania translacji w układzie in vitro. Badano podatność syntetyzowanych białek na działanie proteaz. • mRNA białka wydzielniczego • proteazy degradacja białka • mRNA białka wydzielniczego • proteazy • mikrosomy brak degradacji białka Wynik potwierdzono metodami radioizotopowymi
Jaki mechanizm decyduje o tym czy rybosom przyłączy się do ER czy też nie? • O przyłączeniu się do ER nie decyduje rybosom, lecz samo syntetyzowane białko. • Jeśli porównano sekwencję białek sekrecyjnych produkowanych in vivo i in vitro (bez udziału mikrosomów) okazało się, że białka produkowane in vitro są dłuższe od N-końca niż te natywne.
PEPTYDY SYGNAŁOWE • Peptydy sygnałowe (sekwencje liderowe albo sekwencja „pre”) odpowiadają za oddziaływanie rybosomów syntetyzujących dane białko z ER. • Peptyd sygnałowy wprowadza białko do światła ER; tam ulega odcięciu przez specjalny enzym: peptydazę sygnałową. • Obecnie znanych jest wiele tysięcy sekwencji sygnałowych różnych białek • Cechy peptydu sygnałowego: • Długość: 13 - 36 reszt aminokwasowych. • Zawiera przynajmniej jedną resztę aminokwasową o charakterze zasadowym • (Arg lub Lys), zwykle znajdującą się bardzo blisko N-końca; nawet zaraz po reszcie metioniny. • Zawieraodcinek silnie hydrofobowy (10 -15 reszt: Ala, Leu, Val, Ile, Phe). • Ostatni aminokwas peptydu sygnałowego ma zazwyczaj mały niepolarny łańcuch boczny. Najczęściej jest to alanina.
Przykłady peptydów sygnałowych Albumina Łańcuch lekki IgG Prolaktyna Lizozym Penicylinaza E. coli
N C Sekwencje “pre”, sekwencje “pro” Nie wszystkie białka sekrecyjne mają sekwencję sygnałową (sekwencję “pre”) na N-końcu. Np. owoalbumina ma tę sekwencję w obrębie łańcucha – jest to tzw. wewnętrzna sekwencja sygnałowa (znajduje się i między 22 a 41 aa, nie jest usuwana). Białko posiadające sekwencję sygnałową to PRE-białko. Niektóre białka posiadają również sekwencjęPRO (niektóre hormony, enzymy). Są to Pre-Pro-białka. Sekwencje “pro” mogą występować w różnych miejscach łańcucha polipeptydowego. Utrzymują białko w stanie nieaktywnym biologicznie: proinsulina, trypsynogen, protrombina itd. PRE PRO BIAŁKO
W jaki sposób peptyd sygnałowy odnajduje ER? To wzajemne rozpoznanie zachodzi dzięki parze białek. Są to: SRP(signal recognition particle) – cząstka rozpoznająca sygnał; białko cytoplazmatyczne Receptor SRP – białko występujące w błonie ER SRP - rybonukleoproteina - kompleks o masie 325 kDa (RNA - 300 nukleotydów i 6 różnych łańcuchów polipeptydowych). Gdy peptyd sygnałowy wynurzy się z obrębu rybosomu, odnajduje go białko SRP, które wiąże się zarówno do niego jak i do rybosomu. SRP rozpoznaje peptyd sygnałowy + rybosom. SRP może oddziaływać z samotnym rybosomem ale z niskim powinowactwem. SRP oddziałuje z układem: rybosom/peptyd sygnałowy z wysokim powinowactwem.
SRP jest strażnikiem prawidłowego transportu Doświadczenie: Synteza białka w układzie bezkomórkowym. • podstawowe • składniki układu • SRP • brak mikrosomów zahamowanie translacji • podstawowe • składniki układu • SRP • mikrosomy translacja zachodzi
Jakie znaczenie ma zahamowanie translacji przez SRP przed przyłączeniem się rybosomu do ER? • Zahamowanie translacji przez przyłączenie SRP do układu rybosom/peptyd sygnałowy spełnia dwie funkcje: • Uniemożliwia samorzutne powstanie struktur drugorzędowych i trzeciorzędowych zanim polieptyd nie trafi do ER. • Uniemożliwia zakończenie syntezy i uwolnienie do cytoplazmy białka, które ma być wydzielone albo trafić do lizosomów, ap. Golgiego itp.
RECEPTOR SRP wiąże kompleks SRPpeptyd sygnałowyrybosom Receptor SRP (SRP-R) (białko dokujące) obecny w błonie ER jest zbudowany z dwóch łańcuchów polipeptydowych: (68 kDa) i (30 kDa). Zarówno SRP (jeden z jego łańcuchów polipeptydowych - 54 kDa) jak i obie podjednostki SRP-R są białkami G.
TRANSLOKON Translokon to układ (system), którego funkcją jest przeprowadzenie łańcuchów polipeptydowych przez błonę ER (Eukaryota), lub błonę komórkową(Prokaryota)
NBD – znacznik fluorescencyjny Czas życia fluorescencji: w środowisku hydrofobowym - 7 ns w środowisku hydrofilowym - 1,5 ns Przez cały czas translokacji obserwuje się ten sam, krótki czas fluorescencji (1,5 ns) transport łańcucha polipeptydowego zachodzi przez cały czas w środowisku wodnym istnieją kanały błonowe dla translokacji białka do ER NBD - nitrobenzooksadiozol
TRANSLOKON Translokon tworzy "wodny", (wypełniony cząsteczkami wody), kanał przechodzący przez błonę Głównym składnikiem translokonu jest kompleks 3 białek - podobny u Prokaryota i Eukaryota. U Prokaryota: kompleks SecYEG U ssaków:kompleks Sec61p (3 białka: , , ) Sec61 - zbudowane z 10 homologicznych segmentów przechodzących przez błonę (podobne u Pro- i Eukaryota). Niezbędne dla życia. Sec61 - pojedynczy łańcuch transmembranowy(podobne u Pro- i Eukaryota). Niezbędne dla życia. Sec61 - różne u Pro- i Eukaryota; nie jest niezbędne dla życia. U ssaków współpracują też białka: Sec62, Sec63 oraz TRAM- Translocating chain-Associated Membrane protein (8 x przebijający błonę) TRAP – Translocon Associated Protein complex, heterotetramer
Kanały translokonu • Kanał jest strukturą dynamiczną i wielkość • otworu w obrębie kanału zmienia się. • Średnica poru kanału: • bez związanego rybosomu: 9 - 15 Å • w obecności rybosomu:40 – 60 Å • Kanał jest utworzony przez oligomer • 3 - 4 cząsteczek Sec61. • Średnica 60 Å jest największą wartością • podawaną dla jakiegokolwiek kanału w • błonie biologicznej. • Białka kanału wchodzą w ścisły związek z • rybosomem. Uwaga! Por jądrowy nie jest kanałem. Por jądrowy (sam otwór) ma średnicę od 9 – 26 nm.
http://nobelprize.org/nobel_prizes/medicine/laureates/1999/illpres/protein.htmlhttp://nobelprize.org/nobel_prizes/medicine/laureates/1999/illpres/protein.html
Co warunkuje szczelność translokonu? Doświadczenie:
Jak zostaje zapewniona szczelność ER • Doświadczenie: • Jony jodkowe w zetknięciu z NBD wygaszają jego fluorescencję. • Jeśli doda się jonów jodkowych do układu prowadzącego translację w obecności NBD-Lys i mikrosomów to fluorescencja nie zostaje w ogóle wygaszona połączenie rybosom - ER jest szczelne. • Jeśli teraz “podziurawi” się błonę ERstreptolizyną, jony jodkowe mogą dostać się do łańcucha peptydowego znakowanego NBD od drugiej strony – spodziewamy się wygaszenia fluorescencji. • Ale tak się nie dzieje ..... • aż do momentu, gdy łańcuch polipeptydowy jest dłuższy niż70 aminokwasów.
Nieprzepuszczalność kanału błonowego jest • podwójnie zabezpieczona: • od strony cytoplazmy kanał jest "zapieczętowany" rybososmem • od strony światła ER jest zamknięty przez białko bramkujące • Pozostawienie kanału zamkniętego od strony światła ER, aż do osiągnięcia przez peptyd długości 70 aminokwasów zapewnia nieprzepuszczalność kanału do chwili, gdy oddziaływanie rybosom – translokon będzie właściwe.
Jakie białko pełni funkcjębiałka bramkującego? • Doświadczenie: • Badano wygaszanie fluorescencji NBD-lizyny przez jony jodkowe stosując specjalnie spreparowane mikrosomy. • izolacja mikrosomów • izolacja poszczególnych białek mikrosomów • zamknięcie w mikrosomach po 1 białku • Obserwowano jak takie mikrosomy posiadające tylko wybrane białka są zdolne do blokowania dostępu jonów jodkowych do tworzącego sięłańcucha polipeptydowego. • Wynik: • W mikrosomach zawarte BiP nie ma wygaszania fluorescencji kanał zamknięty. • W mikrosomach brak BiP jest wygaszanie fluorescencji kanał otwarty. • Wniosek: BiP jest białkiem bramkującym.
BiP - początkowo poznane jako • immunoglobulin heavy chain binding protein • Później okazało się, że BiP wiąże w ER wiele białek – jest głównym białkiem opiekuńczym ER. • Jest ATPazą. Aktywność ATPazy (hydroliza ATP) jest wymagana do pełnienia przez BiP zarówno funkcji białka opiekuńczego jak i białka bramkującego. • Inna nazwa Grp78 (glucose-regulated protein 78). Ekspresja wzrasta przy niedoborze glukozy. • Należy do rodziny hsp70. Ekspresja stymulowana przez stres.
Podsumowanie • Receptor SRP oddziałuje z białkami translokonu w łańcuchu b dochodzi do wymiany GDP na GTP zmiana konformacji uwolnienie peptydu sygnałowego do translokonu. • Peptyd sygnałowy wchodzi w interakcję z białkami translokonu. Zaczyna przechodzić przez kanał utworzony przez Sec61. • Równocześnie rybosom wiąże się z translokonem – translokon pełni funkcję receptora rybosomu. • Przez pewien czas światło ER pozostaje zamknięte od wewnątrz przez BiP. Przez ten czas rybosom może się dobrze dopasować do translokonu. Kanał w obrębie rybosomu, w którym znajduje się tworzony łańcuch polipeptydowy umieszczony zostaje bezpośrednio nad kanałem translokonu. • Kanał otwiera się i peptyd wchodzi do światła ER. Równoczesna elongacja i translokacja do ER. • Po zsyntetyzowaniu całego łańcucha i jego wejściu do ER następuje lateralne (poziome) przemieszczenie peptydu sygnałowego z kanału do błony.
wnętrze ER = zewnątrzkomórkowa strona cytoplazmatyczna wnętrze ER = zewnątrzkomórkowa strona cytoplazmatyczna


