Download
1 / 1
10 likes | 161 Views
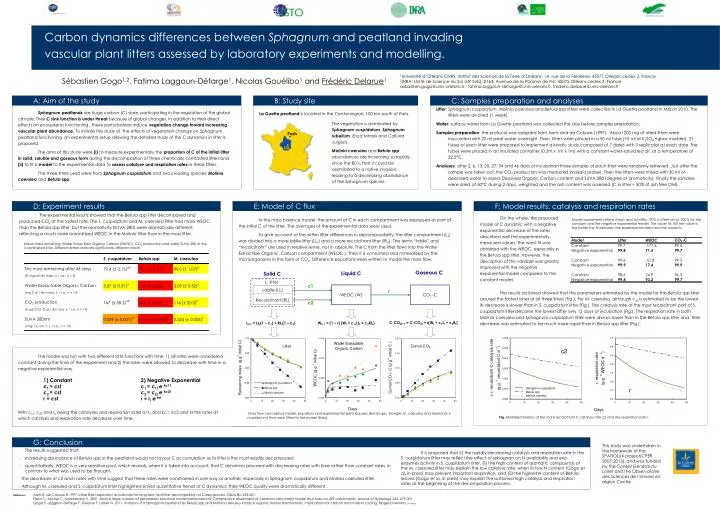
C: Samples preparation and analyses. B: Study site. A: Aim of the study. Paris. LG. L: litter. c1. WEOC (W). CO 2 -C. Labile (LL) Recalcitrant (RL). r. D: Experiment results. c2. F: Model results: catalysis and respiration rates. G: Conclusion.

E N D
C: Samples preparation and analyses B: Study site A: Aim of the study Paris LG L: litter c1 WEOC (W) CO2 -C Labile (LL) Recalcitrant (RL) r D: Experiment results c2 F: Model results: catalysis and respiration rates G: Conclusion Carbon dynamics differences between Sphagnum and peatland invading vascular plant litters assessed by laboratory experiments and modelling. Sébastien Gogo1,2, Fatima Laggoun-Défarge1, Nicolas Gouélibo1 and Frédéric Delarue1 1Université d’Orléans-CNRS, Institut des Science de la Terre d’Orléans, 1A, rue de la Férollerie, 45071 Orléans cedex 2, France 2INRA- Unité de Science du Sol (UR Sols), 2163, Avenue de la Pomme de Pin, 45075 Orléans cedex 2, France sebastien.gogo@univ-orleans.fr , fatima.laggoun-defarge@cnrs-orleans.fr, frederic.delarue@univ-orleans.fr Litter: Sphagnum cuspidatum, Molinia caerulea and Betula spp litters were collected in La Guette peatland in March 2010. The litters were air-dried (1 week) Water: surface water from La Guette peatland was collected the day before samples preparation. Samples preparation: the protocol was adapted from Aerts and de Caluwe (1997). About 200 mg of dried litters were inoculated with 20 ml peat water overnight. Then, litters were placed in a 50 ml tube (+5 ml of K2SO4 +glass marbles). 21 tubes of each litter were prepared to implement a kinetic study composed of 7 dates with 3 replicates at each date. The tubes were placed in an insulated container (0.3m x 1m x 1m) with a constant water-saturated air, at a temperature of 22.5°C. Analyses: after 2, 6, 13, 20, 27, 34 and 46 days of incubation three samples of each litter were randomly retrieved. Just after the sample was taken out, the CO2 production was measured (Vaisala probe). Then the litters were rinsed with 30 ml of deionised water to assess Dissolved Organic Carbon content and SUVA 280 (degree of aromaticity). Finally the samples were dried at 50°C during 2 days, weighted and the ash content was assessed (C in litter = 50% of ash free OM). Sphagnum peatlands are huge carbon (C) store, participating in the regulation of the global climate. Their C sink function is under threat because of global changes. In addition to their direct effects on ecosystems functioning , these perturbations induce vegetation change toward increasing vascular plant abundance. To initiate the study of the effects of vegetation change on Sphagnum peatland functioning, an experimental setup allowing the detailed study of the C dynamics in litter is proposed. The aims of this study were (i) to measure experimentally the proportion of C of the initial litter in solid, soluble and gaseous form during the decomposition of three chemically contrasted litters and (ii) to fit a model to the experimental data to assess catalyse and respiration rates in these litters. The three litters used were from Sphagnum cuspidatum and two invading species: Molinia caerulea and Betula spp. La Guette peatland is located in the Centre region, 100 km south of Paris. The vegetation is dominated by Sphagnum cuspidatum, Sphagnum rubellum, Erica tetralix and Calluna vulgaris. Molinia caerulea and Betula spp abundances are increasing so rapidly since the 80’s, that it could be assimilated to a native invasion, leading to a decreasing abundance of the Sphagnum species. E: Model of C flux The experimental results showed that the Betula spp litter decomposed and produced CO2 at the fastest rate. The S. cuspidatum and M. caerulea litter had more WEOC than the Betula spp litter, but the aromaticity (SUVA 280) were dramatically different, reflecting a much more aromatized WEOC in the Molinia litter than in the moss litter. On the whole, the proposed model of C dynamic with a negative exponential decrease of the rates described well the experimentally measured values. The worst fit was obtained with the WEOC, especially in the Betula spp litter. However, the description of this variable was greatly improved with the negative exponential model compared to the constant model. In the mass balance model, the amount of C in each compartment was expressed as part of the initial C of the litter. The averages of the experimental data were used. To give account of the within litter differences in decomposability, the litter compartment (Lt) was divided into a more labile litter (LLt) and a more recalcitrant litter (RLt). The terms “labile” and “recalcitrant” are used in relative sense, not in absolute. The C from the litter flows into the Water Extractible Organic Carbon compartment (WEOC ). Then it is consumed and mineralized by the microorganisms in the form of CO2. Difference equations were written to model the mass flow. Model assessment criteria (Nash and Sutcliffe, 1970, in Perrin et al. 2001) for the constant and the negative exponential model. The closer to 100 the value is, the better the fit between the experimental data and the model is. Mean mass remaining, Water Extractible Organic Carbon (WEOC), CO2 production and water SUVA 280 of the 3 contrasted litter. Different letters indicate significantly different mean. Gaseous C Liquid C Solid C The results obtained showed that the parameters estimated by the model for the Betula spp litter assured the fastest rates of all three litters (Fig.). For M. caerulea, although c20 is estimated to be the lowest, its decrease is slower than in S. cuspidatum litter (Fig.). The catalysis rate of the most recalcitrant part of S. cuspidatum litter became the lowest after only 12 days of incubation (Fig.). The respiration rate in both Molinia caerulea and Sphagnum cuspidatum litter were always lower than in the Betula spp litter and, their decrease was estimated to be much more rapid than in Betula spp litter (Fig.). C-CO2t+1 = C-CO2t + r(Wt + c1Lt + c2Rt) Lt+1 = LLt(1 – c1) + RLt(1 – c2) Wt+1 = (1 – r) (Wt + c1LLLt + c2RLt) The model was run with two different rate functions with time: 1) all rates were considered constant during the time of the experiment and 2) the rates were allowed to decrease with time in a negative exponential way. With c10, c20 and r0 being the catalyses and respiration rates at t0 and kc1, kc2 and kr the rates at which catalysis and respiration rate decrease over time. 1) Constant c1 = cst c2 = cst r = cst 2) Negative Exponential c1 = c1i e-kc1t c2 = c2i e-kc2t r = ri e-krt Mass flow conceptual model, equations and experimental data (square; Betula spp, triangle: M. caerulea and diamond: S. cuspidatum) that were fitted to the model (lines). Fig. Modelled kinetics of the more recalcitrant C catalysis rate c2 and the respiration rate r. This study was undertaken in the framework of the SPATIOLUX project(CPER 2007-2013), and was funded by the Conseil Général du Loiret and the Observatoire des Sciences de l’Univers en région Centre • The results suggested that: • Increasing abundance of Betula spp in the peatland would not favour C accumulation as its litter is the most readily decomposed • quantitatively, WEOC is a very sensitive pool, which reveals, when it is taken into account, that C dynamics proceed with decreasing rates with time rather than constant rates, in contrary to what was used to be thought, • - the decreases of c2 and r rates with time suggest that these rates were constrained in one way or another, especially in Sphagnum cuspidatum and Molinia caerulea litter, • - Although M. caerulea and S. cuspidatum litter highlighted similar quantitative trends of C dynamics, their WEOC quality were dramatically different . It is proposed that (i) the rapidly decreasing catalysis and respiration rate in the S. cuspidatum litter may reflect the effect of sphagnan on N availability and exo-enzymes activity in S. cuspidatum litter, (ii) the high content of aromatic compounds of the M. caerulea litter may explain the low catalysis rate, when its low N content (Gogo et al, in press) may prevent important respiration, and (iii) the highest N content of Betula leaves (Gogo et la, in press) may explain the sustained high catalysis and respiration rates at the beginning of the decomposition process. Référence: Aerts R, de Caluwe H.1997. Initial litter respiration as indicator for long-term leaf litter decomposition of Carex species. Oikos 80: 353-361. Perrin C, Michel C, Andréassian V. 2001. Does a large number of parameters enhance model performance? Comparative assessment of common catchment model structures on 429 catchments. Journal of Hydrology 242: 275-301. Gogo S, Laggoun-Défarge F, Delarue F, Lottier N. 2011. Invasion of a Sphagnum-peatland by Betula spp and Molinia caerulea impacts organic matter biochemistry. Implications for carbon and nutrient cycling. Biogeochemistry, in press.








