Download
1 / 27
290 likes | 575 Views
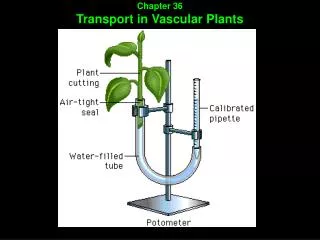
Transport in vascular plants includes: 1 - transport of water and solutes by individual cells 2 - short-distance transport of substances from cell to cell 3 - long-distance transport within xylem and phloem Figure #1 (right)

E N D
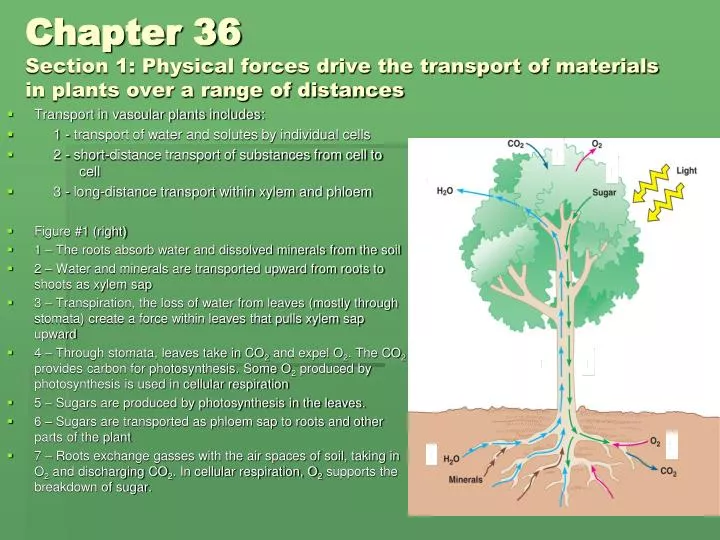
Transport in vascular plants includes: 1 - transport of water and solutes by individual cells 2 - short-distance transport of substances from cell to cell 3 - long-distance transport within xylem and phloem Figure #1 (right) 1 – The roots absorb water and dissolved minerals from the soil 2 – Water and minerals are transported upward from roots to shoots as xylem sap 3 – Transpiration, the loss of water from leaves (mostly through stomata) create a force within leaves that pulls xylem sap upward 4 – Through stomata, leaves take in CO2 and expel O2. The CO2 provides carbon for photosynthesis. Some O2 produced by photosynthesis is used in cellular respiration 5 – Sugars are produced by photosynthesis in the leaves. 6 – Sugars are transported as phloem sap to roots and other parts of the plant 7 – Roots exchange gasses with the air spaces of soil, taking in O2 and discharging CO2. In cellular respiration, O2 supports the breakdown of sugar. Chapter 36Section 1: Physical forces drive the transport of materials in plants over a range of distances
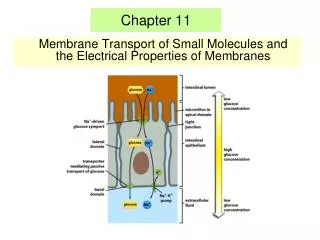
Selective Permeability of Membranes: A Review • passive transport- diffusion across a membrane down a gradient; happens without the cell directly using metabolic energy • active transport- the pumping of solutes across membranes against their electrochemical gradients, the combined effects of the concentration gradient and the voltage (charge difference) across the membrane; the cell must expend energy, usually in the form of ATP • transport proteins- embedded in the membrane; help solutes cross the lipid bilayer of the membrane; may bind selectively to a solute on one side of a membrane and release it on the opposite side, or they may provide a selective channel across the membrane
The Central Role of Proton Pumps • proton pump- uses energy from ATP to pump hydrogen ions (H+) out of the cell • The pumping of hydrogen ions out of the cell results in a proton gradient with a higher H+ concentration outside the cell than inside. The hydrogen ions diffuse back into the cell. This “flow” can be harnessed as energy to do work. • membrane potential- a separation of opposite charges across a membrane. Charge separation is a form of potential energy that can be harnessed to perform cellular work. • Proton pumps contribute to membrane potential. • In plant cells, energy stored in the proton gradient and • membrane potential to drive the transport of many • different solutes. • cotransport- a transport protein couples the downhill • passage of one solute to the uphill passage of another. • Cotransport is responsible for the uptake of sucrose by • plant cells. • chemiosmosis- uses proton pumps; has a • transmembrane proton gradient that links energy- • releasing processes to energy-consuming processes • in cells.
Effects of Differences in Water Potential • osmosis- the passive transport of water across a membrane • The net uptake or loss of water by a cell occurs by osmosis. • Water moves by osmosis from the solution with a lower solute concentration to the solution with a higher solute concentration. • Physical pressure is also a factor in osmosis. • water potential (abbreviated )- the combined effects of solute concentration and physical pressure; determines the direction of movement of water • Free water (water that is not bound to solutes or surfaces) moves from regions of higher water potential to regions of lower water potential if there is no barrier to its flow. • megapascals (abbreviated MPa)- a unit of pressure used to measure water potential.
Effects of Differences in Water Potential: continuedHow Solutes and Pressure Affect Water • solute potential (abbreviated S)- proportional to the number of dissolved solute molecules. • Solute potential is also called osmotic potential because solutes affect the direction of osmosis. • pressure potential- the physical pressure on a solution. It can be positive or negative relative to atmospheric pressure • tugor pressure- produced when cell contents press the plasma membrane against the cell wall
Effects of Differences in Water Potential: continuedQuantitative Analysis of Water Potential • In the absence of physical pressure (P = 0) the water potential () will be equal to the solute potential (S), and the water will flow towards the region of low solute potential (the region with more dissolved solute molecules). • If pressure equal to the positive value of the negative solute potential is applied to the region with the dissolved solute, the water potential would equal zero, and the water would be at equilibrium if the other side of the membrane contained no dissolved solutes (= 0). • If pressure greater than the positive value of the negative solute potential is applied to the region with the dissolved solute, the water potential would be greater than zero, and if the other side of the membrane contained no dissolved solutes (= 0), water would flow towards the side with no dissolved solutes. • If pressure was lessened on the side with no dissolved solute molecules to less than the solute potential of the side with dissolved solutes, the water would flow towards the side with no dissolved solutes.
Quantitative Analysis of Water Potential: Continued • flaccid- limp; a cell that is flaccid has a water potential of 0. • plasmolyze- occurs when the cell’s protoplast shrinks and pulls away from the wall; This happens when the cell has a higher water potential than the outside of the cell. Water then leaves the cell and the cell plasmolyzes. • turgid- very firm; occurs when the water potential of a cell is lower than the water potential outside the cell. Healthy cells are turgid. turgor contributes to support. • wilting- the drooping of leaves and stems as a result of cells becoming flaccid
Effects of Differences in Water Potential: continuedAquaporin Proteins and Water Transport • aquaporins- transport proteins that help water cross vacuolar and plasma membranes • Aquaporins control the rate at which water diffuses down its water potential gradient. They do not affect the water potential gradient or the direction of water flow. • Rate of water movement through aquaporins may be regulated by phosphorylation of the aqua Orin proteins induced by changes in second messengers.
Three Major Compartments of Vacuolated Plant Cells • Transport is regulated by the compartmental structure of plant cells. The plasma membrane functions as a selectively permeable membrane that controls the traffic of molecules into and out of the protoplast. It is a barrier between the cell wall and the cytosol, two major compartments of the cell. • The vacuole is a large organelle that can occupy as much as 90% or more of the protoplast’s volume. • vacuolar membrane or tonoplast- regulates molecular traffic between the cytosol and vacuolar contents (cell sap) • Cell walls and cytosol are continuous from cell to cell. They are connected by plasmodesmata. • symplast- the cytoplasm continuum • apoplast- the continuum of cell walls plus the extra cellular spaces • The vacuole is the third cellular compartment, and is not shared with neighboring cells
Functions of the Symplast and Apoplast in Transport • Short-distance transport, such as from the root hairs to the vascular cylinder of the root, is called lateral transport because it is usually along the radial axis of plant organs, rather than up and down along the length of the plant. • Lateral transport may be accomplished by three different routes. • 1 - transmembrane route - substances move out of one cell, across the cell wall, and into the next cell, which may then pass the substance along to the next cell • 2 - via symplast - requires only one crossing of a plasma membrane; After entering one cell, solutes and water move from cell to cell via plasmodesmata • 3 - along the apoplast - water and solutes move from one location to another within a root or other organ along the byways provided by the continuum of cell walls; the solute never enters a protoplast
Bulk Flow in Long-Distance Transport • bulk flow- the movement of a fluid driven by pressure; contributes to long-distance transport • In bulk flow, water and solutes moves through the tracheids and vessels of the xylem and through the sieve tubes of the phloem. • In phloem, the loading of sugar generates a high positive pressure at one end of a sieve tube, forcing sap to the opposite end of the tube. • Xylem use tension (negative pressure) to drive long distance transport • Transpiration is the evaporation of water from a leaf. It reduces pressure in the leaf xylem, creating tension that pulls xylem sap upward from the roots.
Chapter 36Section 2: Roots absorb Water and Minerals from the soil • In the Apoplastic route, the hydrophilic walls of the root hairs take in the water and minerals from the soil, providing access to the apoplast. • In the Symplastic route, the water and minerals enter the symplast through the plasma membrane of the root hairs. • During the Apoplastic route, some water and soil enter the protoplasts of cells through the symplast when passing trough the epidermis and the cortex. • Endodermal cells contain a casparian strip (shown here in purple) that does not allow water or minerals to pass. In order to overcome this barrier, the material must enter the symplast ,if not already there, to get into the vascular cylinder. • Endodermal and parenchyma cells discarche water and mineralsoinot the apoplast, allowing the xylem vessels to receive it and transport it to the shoot system.
The Roles of Root Hairs, Mycorrhizae, and Cortical Cells • Root hairs are extensions of epidermal cells that allow soil particles coated with water and minerals to adhere to them. Once the soil solution is absorbed by the hydrophilic walls, it is exposed to the symplast of all the epidermal cells and the cortical cells. • As material is being absorbed, roots can accumulate vital minerals in concentrations hundreds of times higher than the soil. • Mycorrhizea are symbiotic structures formed by roots and fungi. These structures allow an increased surface area for water and select materials to be absorbed.
The Endodermis: A Selective Sentry • The Endodermis is located at the end of the cortex. It serves as a checkpoint before material is transmitted throughout the rest of the plant. Minerals traveling through the symplastic route pass easily through the endodermis, but those passing through the apaplastic route must enter they symplast. This happens because endodermal cells contain the Casparian strip. • The Casparian strip is made of suberin which does not allow the passage of water or any other minerals. This strip assures that only selected minerals can pass through to the xylem vessels and solutes that have accumulated in the xylem sap do not flow back into the soil solution. • Once the minerals pass the Casparian strip, endodermal cells and parenchyma cells discharge the minerals from the symplast to the apoplast. This lets the material enter the tracheids and vessel elements of the xylem. These are water conducting and part of the apoplast since they do not contain a protoplast when mature. These parts make up the end of the soil to xylem pathway and the material can now be passed to the shoot system.
Chapter 36 Section 3: Water and minerals ascend from roots to shoots through the xylem • Leaves depends on the long-distance transport system of xylem sap which is flowing upward from roots throughout the shoot system to veins that branch throughout each leaf for their supply of water and mineral nutrients. • Transpiration is the loss of astonishing amount of water vapor from leaves usually through stomata, the microscopic pores on the surface of a leaf and other aerial parts of the plant by diffusion and evaporation. • When the transpiration is very low or zero, usually at night, the root cells continue to pump mineral ions into the xylem of the vascular cylinder. At the same time endodermis helps prevent the ions from leaking out. This accumulation of minerals lowers the water potential within the vascular cylinder. • Factors affecting the Ascent of Xylem sap -Root pressure is an upward push of xylem sap which allows water to flow in from the root cortex and cause guttation by making more water to enter the leaves than is transpired. -Guttationis the exudation of water droplets that can be seen in the morning on the tips of grass blades or the leaf margins of some small, herbaceous eudicots. It is different from dew. (figure 34.11) -In most of plants, root pressure is a minor mechanism driving the ascent of xylem sap, at most forcing water upward only a few meters.
The generation of transpirational pull in a leaf • 1) In transpiration, water vapor (shown as blue dots) diffuses from the moist air spaces of the leaf to the drier air outside via stomata. • 2) At first, the water vapor lost by transpiration is replaced by evaporation from the water film that coats mesophyll cells. • 3) Evaporation causes the air-water interface to retreat farther into the cell wall and become more curved as the rate of transpiration increases. As the interface becomes more curved, the water film’s pressure becomes more negative. This negative pressure, or tension, pulls water from the xylem, where the pressure is greater.
Water and minerals ascend from roots to shoots through the xylem • The transpiration-cohesion-tension mechanism is responsible for pulling or transporting xylem sap against its gravity: -Transpiration pull: When the transpiration occurs severely, the degree of curvature and the surface tension of the water molecules increases, accordingly the pressure at the air-water interface becomes increasingly negative and more water molecules are pulled toward this region to reduce the tension. *Negative pressure (tension) lowers water potential and thus, it causes the “pull” in transpirational pull. -Cohesion and Adhesion facilitate the long-distance transport throughout the plant *The cohesion of water due to hydrogen bonding between the water molecules makes it possible to pull a column of sap from above without the water molecules separating. *The strong adhesion of water molecules (again by hydrogen bonds) to the hydrophilic walls of xylem cells aids in offsetting the downward pull of gravity. *Some other plant structures, such as the thick secondary walls help to prevent vessels from collapsing (structural support). * Transpirational pull can extend down to the roots only through an unbroken chain of water molecules -Capitation is the formation of a water vapor pocket in a vessel breaks the chain; the air bubbles resulting from cavitations expand and eventually become embolisms, blockages of the water channels of the xylem. -Root pressure enable small plants to refill embolized vessels in spring ; Only youngest, outermost secondary xylem transports water while the older secondary xylem does provide support for the tree.
Water and minerals ascend from roots to shoots through the xylem • Xylem Sap Ascent by Bulk Flow : In the long-distance transport of water from roots to leaves by bulk-flow, the movement of fluid is driven by a water potential difference at opposite ends of a conduit and it is only depending on pressure. (In case of plants, the conduits are vessels or chains of tracheids.) -The plant absorbs the sunlight to drive transpiration by causing water to evaporate from the moist walls of mesophyll cells and by lowering the water potential in the air spaces within a leaf rather than expending any type of energy to lift xylem sap by bulk flow. - Unlike osmosis, which moves only water, the bulk flow moves the whole solution, water plus minerals and any other solutes dissolved in the water. • The water potential difference is generated at the leaf end by transpirational pull, which lowers the potential (increase tension) at the “upstream” end of the xylem. -Water potential gradients drive the osmotic movement of water from cell to cell within root and leaf tissue ; * Difference in both solute concentration and turgor pressure contribute to this short-distance transport.
Chapter 36: section4: Stomata help regulate the rate of transpiration • Leaves generally have large surface areasand high surface area-to-volume ratios. • Large surface area morphological adaptation that enhances the absorption the absorption of light needed to drive photosynthesis • High surface area-to-volume ratio aids in the uptake of carbon dioxide during photosynthesis and well as the release of oxygen produces as a by-product of photosynthesis • Large surface areas and high surface area-to-volume ratios have the serious drawback of increasing water loss by way of the stomata. • Guard cells help balance the plant’s requirements to conserve water with its requirement for photosynthesis.
Effects of Transpiration on Wilting and Leaf Temperature • A leaf may transpire more than its weight in water each day, and water may move through the xylem at a rate as fast as 75 cm/min. • If transpiration continues to pull sufficient water upward to the leaves, they will not wilt. • Some evaporative water loss does occur even when the stomata are closed. • Transpiration also results in evaporative cooling, which can lower the temperature of a leaf by as much as 10-15C compared with the surrounding air.
Stomata: Major Pathways for Water Loss • About 90% of the water a plant loses escapes through stomata, though these pores account for only 1-2% of the external leaf surface. • Cuticle limits water loss through the remaining surface of the leaf • Guard cells control the diameter of the stoma by changing shape, thereby widening or narrowing the gap between the two cells. • The stomatal density of a leaf is under both genetic and environmental control. • High light intensities and low carbon dioxide levels during leaf development tend to increase stomatal density in many plant species. • When guard cells take in water from neighboring cells by osmosis, they become more turgid and bowed. • The changes in turgor pressure that open and lose stomata results primarily from the reversible uptake and loss of potassium ions by the guard cells. • The K+ fluxes across the guard cell membrane are coupled to the generation of membrane potentials by proton pumps. • In general, stomata are open during the day and closed during the night. • This prevents the plant from losing water when it is too dark for photosynthesis. • A second stimulus causing stomata to open is depletion of carbon dioxide within air spaces for the leaf, which occurs when photosynthesis begins in the mesophyll. • A third cue causing stomatal opening is an internal “clock” in the guard cells. • Cycles that have intervals of appx. 24 hours are called circadian rhythms. • Environmental stresses can cause stomata to close during the day time. • Guard cells arbitrate the photosynthesis-transpiration compromise on a moment-to-moment basis by integrating a variety of internal and external stimuli.
Xerophyte Adaptations that reduce transpiration • Plants adapted to arid climates, called xerophytes, have various leaf modifications that reduce the rate of transpiration. • Many xerophytes have small, thick leaves, an adaptation that limits water loss by reducing surface area relative to leaf volume. • An elegant adaptation to arid habitats is found in succulents of the family Crassulaceae, in ice plants, and in many other plant families. • These plants assimilate carbon dioxide by an alternative photosynthetic pathway known as CAM, for crassulacean acid metabolism
Chapter 36section 5: Organic nutrients are translocated through the phloem Translocation- the transport of organic nutrients in a plant In order to transport sugars from leaves to other parts of the plant xylem sap flows from roots to leaves in the opposite direction Products of photosynthesis are transported via the phloem, which is a second vascular tissues
Movement from Sugar Sources to sugar sinks • Sieve tube members are the specialized cells of phloem that function as the conduits for translocation in angiosperms, these members when placed together from sieve tubes. • Between the cells are sieve plates which allow the flow of sap along the sieve tube. • Phloem sap is an aqueos solution that is different from xylem sap, it is primarily made up of dissacharide sucrose. The sucrose concentration can be up to 30% and gives the sap its thickness. • Sieve tubes carry sugar from the sugar source to the sugar sink Sugar source – a plant organ that is the net producer of sugar Sugar sink – a plant organ that is the net consumer of sugar • Some spieces move sugar from the mesophyll to the sieve tube via symplast, which means passing the plasmodesmata. Then it moves into the apoplast and is then accumulated by the sieve tube • Transfer cells- cells that enhance the transfer of solutes between apoplast and symplast • The sugar level in the sink is always lower, because it is either consumed during growth or converted into another polymer
Pressure Flow: The Mechanism of Translocation in Angiosperms • Phloem sap flows at a rate of 1 m/hr • When scientist studied angiosperms, they determined that sap flows through the sieve tube in bulk flow, and is driven by pressure flow • Sugar transport can be seen on three levels • Cellular level across the plasmadesmata- sucrose accumulation via active transport • Short-distance transport –sucrose migration from the mesophyll to the phloem • Long distance transport between organs – bulk flow in sieve tubes.