Download

1 / 24
300 likes | 480 Views
Conserved ligands, cell-surface receptors and signaling co-receptors from C. elegans to humans 3 subfamilies: TGF-b, activin/inhibin/nodal, BMP Ligands are both homodimers or heterodimers with a set of 7 cysteines: 3 intrasubunit disulfide bonds + 1 intersubunit bond

E N D
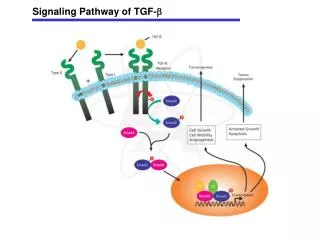
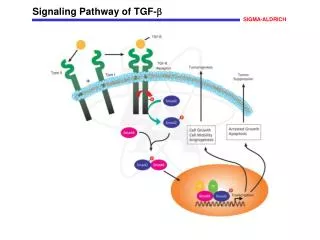
Conserved ligands, cell-surface receptors and signaling co-receptors from C. elegans to humans 3 subfamilies: TGF-b, activin/inhibin/nodal, BMP Ligands are both homodimers or heterodimers with a set of 7 cysteines: 3 intrasubunit disulfide bonds + 1 intersubunit bond Two types cell surface receptors: serine-threonine kinases= type I and type II Receptors All type I receptors have a TTSGSGSG motif, called the GS domain 4 mammalian type II receptors: constitutively active 7 type I receptors= activin receptor like kinases 1-7 (ALK1-7) A TGF ligand signals by binding to and bringing together type I and type II receptors to form a ternary holo-complex. The assembly dynamic differs between ligands In the holo-complex, constitutively active type II kinase transphosphorylates the GS domain of type I, which then phosphorylates Smads 8 Smads, with 3 functional classes: receptor regulated Smads=R-Smad (1,5,8//2,3), co-mediator Smad= co-Smad (4), inhibitory Smad=I-Smad(6,7) TGF-b Superfamily
TGF-b Superfamily 8 Smads, with 3 functional classes: receptor regulated Smads=R-Smad (1,5,8//2,3), co-mediator Smad= co-Smad (4), inhibitory Smad=I-Smad(6,7) After phopshorylation, 2 or 3 activated R-Smad di-tri-merize and form heterometric complexes with smad4.--->nucleus the I-Smads negatively regulate the signaling by competing with R-Smads for the receptors or co-Smad interaction and by targeting the receptors for degradation Signaling through non-Smad pathways: MAPK and ERK, p38, JNK, PI3K/Akt pathway, NF-kB pathway
TGF-b signalling is regulated at all levels TGF-bs are secreted as inactive precursor proteins BMPs are secreted in their active form, but activity is regulated by ligand antagonists: 3 families of BMP antagonists based on the size of their cysteine knot TGF-b co-receptors: -enhance ligand-binding to the receptors -antagonize ligand function by shedding of extracellular domains -regulate receptor localization and internalization -regulate cell adhesion -regulate signalling Pseudo-Receptor FK506-binding protein 12 (FKBP12), binds the GS domain of type I receptors and maintains them inactive. Localization and trafficking of the signaling receptors: SARA or co-receptors. TGF-b receptor trafficking and turnover
Extracellular Antagonists and Membrane Level Regulation Noggin:BMP-7 complex Follistatin:activin complex
TGF-b activation The 3 TGF-b isoforms are synthesized and homodimeric pro-TGF-b = TGF covalently linked to latency associated protein (LAP) Intracellularly cleaved by furin-type enzymes--> mature TGF-b , remains non-covalently associated with LAP= SLC Few cells produce and secrete diffusible SLC. Majority secrete TGF as part of the LLC, formed within secretory vesicles: covalent disulfide bond between the SLC and a member of the LTBP family LTBP-1,-3,-4 bind TGF-bs. LTBPs belong to the superfamily of fibrillin-like ECM proteins Bind a series of ECM proteins TGF-b is synthesized in excess, and bioavailability depends on activation
Activation of TGF-b varies according to cell type and physiological context Heat, acid, reactive oxygen species, glycosidases, thrombospondin, proteolysis and integrin- mediated activation Proteases: Plasmin, thrombin, elastase, MT-MMP,MMP-2, MMP3, MMP-9, MMP13 etc.. in vitro. In vivo, however, simple proteolysis has not been shown. No supporting genetic evidence.. Predominantly non-proteolytic mechanism for TGF-b activation in vivo: Integrin (av, b6, b8) and TGF-b KO mice have overlapping phenotypes. The TGF-b1 RGD->RGE mutation phenocopies the TGF-b1 KO. Integrins shown to bind and possibly activate latent TGF-b1 avb1: binding yes, activation not known. avb3: binding yes, activation yes. avb5: binding yes, activation yes. avb6: binding yes, activation yes. avb8: binding yes, activation yes. a8b1: binding yes, activation no xb1: binding yes, activation yes. Latent TGF-b activation
Integrins bind to the RGD sequence at the N-term of LAP. Mutation to RGE abolishes binding and activation in epithelial cells RGD peptides reduce TGF-b activation in myofibroblasts The TGF-b1 RGD->RGE mutation phenocopies the TGF-b1 KO. avb6 has a higher affinity for the RGD in LAP than in FN due to a DLXXL motif. Some LTBPs also have an RGD, but it has not been shown to be an integrin ligand. Why do not all integrins that bind to the RGD in LAP activate TGF-b1?? Integrins shown to bind and possibly activate latent TGF-b1 avb1: binding yes, activation not known. avb3: binding yes, activation yes. avb5: binding yes, activation yes. avb6: binding yes, activation yes. avb8: binding yes, activation yes. a8b1: binding yes, activation no xb1: binding yes, activation yes. Latent TGF-b activation
Latent TGF-b activation: integrin dependent proteolysis Incubation of purified integrins with TGF-b is not sufficient to activate the latent complex. Mechanism A: integrin docks latent TGF-b and a protease and brings them close together avb8 said to mediate LAP-b1 activation in conjunction with MT-1 MMP: MMP inhibitor + co-localization avb3 activation coincides with MMP-9 and MMP-2 production in cancer cells Direct interaction of avb3 and TGF-b-RII upon TGF-b application has been shown by BRET
Latent TGF-b activation: integrin-mediated cell traction Mechanism B: traction forces due to integrin binding to latent TGF-b results in a conformational change that liberates TGF-b. avb6, avb5, and possibly avb3 can activate Latent TGF-b independently of proteases LAP-b1 has 2 conformations (far UV dichroism spectroscopy): only 1 binds to TGF-b1 There needs to be An actin cytoskeleton Specific integrins Incorporation of LLC into the ECM A mechanically resistant ECM
Latent TGF-b activation: integrin-mediated cell traction • A)Direct activation of latent TGF-b by integrins require a contractile cytoskeleton. • avb6: disruption of actin cytoskeleton with cytochalasin D or cytoplasmic truncation prevents latent TGF-b activation • Mefs with constitutively active RhoA activate latent TGF-b more. (less with dominant neg) • Highly contractile fibroblast but not low contractile fibroblasts release active TGF-b from the same LLC-rich ECM. • Inducing myofibroblast contraction with thrombin, angiotensin-II and endothelin-I increases TGF-b activation: integrin-dependant and protease-independant • Myofibroblast contraction blockers (ML-7, blebbistatin, cytochalasin, SMA-FP) inhibits TGF-b activation by contraction-inducers • External stretch of the myofibroblasts also activates latent TGF-b activation • C)LAP-b1 must be associated with LTBP in the LLC and be incorporated into the ECM for activation of TGF-b by integrins • Deletion of the ECM-binding hinge region of LTBP-1 (or substitution by LTBP-3 which lacks the hinge) abolishes avb6 integrin-mediated activation of latent TGF-b • Expression of an engineered LTBP with only the ECM-binding hinge region and the LAP binding domain is sufficient to promote avb6 integrin-mediated activation of latent TGF-b • different LTBP isoforms possibly regulate latent TGF-b activation • Does the matrix composition do as well???
Latent TGF-b activation: integrin-mediated cell traction D)the efficiency of latent TGF-b activation by integrins depends on the mechanical properties of the ECM Tissue culture dish=rigid substrate Lung tissue of living mice: PAR1-activating peptides are not sufficient for TGF-b activation. Lungs must be mechanically challenged by hight-tidal-volume ventilation+ PAR1-activating peptides Myofibroblasts activate latent TGF-b by integrin-mediated contraction when cultured on stiff silicone substrates, but not on silicone substrates that have the compliance of normal connective tissue.
Latent TGF-b activation Activation of TGF-b varies according to cell type and physiological context Heat, acid, reactive oxygen species, proteolysis and integrin mediated activation Proteases: Plasmin, thrombin, elastase, MMP-2, MMP-9 in vitro. In vivo,
TGF-b and Angiogenesis Loss of TGFb component->abnormal differentiation and maturation of primitive vascular plexus TGFb1-> 50% E9-E10 lethality due to defects in yolk sac vasculature and hematopoietic system. Vessels with decreased wall integrity TGFbRII, ALK5, ALK1, endoglin and Smad5 ->idem However, TGFb1 and TGFbRII are critical for formation of the vascular plexus and remodeling into a complex network. ALK1, ALK5 and Eng only for angiogenesis Endolethial-specific KO of ALK5 and TGFbRII phenocopy the complete KOs
TGF-b and Angiogenesis Tgfb1 and TGFb3
TGF-b and Angiogenesis TGF-b1 has bifunctional effects on endothelial cells in vitro. It can both stimulate and inhibit proliferation