Download
1 / 1
10 likes | 113 Views
b. a. d. f. e. Fall 2010 GSA Annual Meeting Denver, CO Impacts of Land Use and Climate Change on Water Resources Sustainability Session . PHYSICAL PROCESSES AFFECTING THE DISTRIBUTION OF Didymosphenia geminata BLOOMS IN RAPID CREEK, SD-part-I.

E N D
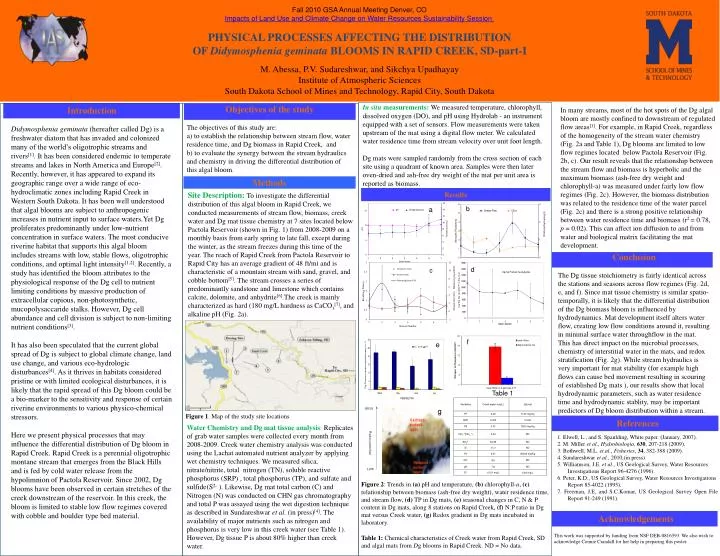
b a d f e Fall 2010 GSA Annual Meeting Denver, CO Impacts of Land Use and Climate Change on Water Resources Sustainability Session PHYSICAL PROCESSES AFFECTING THE DISTRIBUTION OF Didymosphenia geminata BLOOMS IN RAPID CREEK, SD-part-I M. Abessa, P.V. Sudareshwar, and Sikchya Upadhayay Institute of Atmospheric Sciences South Dakota School of Mines and Technology, Rapid City, South Dakota In situ measurements: We measured temperature, chlorophyll, dissolved oxygen (DO), and pH using Hydrolab - an instrument equipped with a set of sensors. Flow measurements were taken upstream of the mat using a digital flow meter. We calculated water residence time from stream velocity over unit foot length. Dg mats were sampled randomly from the cross section of each site using a quadrant of known area. Samples were then later oven-dried and ash-free dry weight of the mat per unit area is reported as biomass. Objectives of the study The objectives of this study are: to establish the relationship between stream flow, water residence time, and Dg biomass in Rapid Creek, and to evaluate the synergy between the stream hydraulics and chemistry in driving the differential distribution of this algal bloom. Introduction Didymosphenia geminata (hereafter called Dg) is a freshwater diatom that has invaded and colonized many of the world’s oligotrophic streams and rivers[1]. It has been considered endemic to temperate streams and lakes in North America and Europe[2]. Recently, however, it has appeared to expand its geographic range over a wide range of eco-hydroclimatic zones including Rapid Creek in Western South Dakota. It has been well understood that algal blooms are subject to anthropogenic increases in nutrient input to surface waters.Yet Dg proliferates predominantly under low-nutrient concentration in surface waters. The most conducive riverine habitat that supports this algal bloom includes streams with low, stable flows, oligotrophic conditions, and optimal light intensity[1,2]. Recently, a study has identified the bloom attributes to the physiological response of the Dg cell to nutrient limiting conditions by massive production of extracellular copious, non-photosynthetic, mucopolysaccaride stalks. However, Dg cell abundance and cell division is subject to non-limiting nutrient conditions[3]. It has also been speculated that the current global spread of Dg is subject to global climate change, land use change, and various eco-hydrologic disturbances[4]. As it thrives in habitats considered pristine or with limited ecological disturbances, it is likely that the rapid spread of this Dg bloom could be a bio-marker to the sensitivity and response of certain riverine environments to various physico-chemical stressors. Here we present physical processes that may influence the differential distribution of Dg bloom in Rapid Creek. Rapid Creek is a perennial oligotrophic montane stream that emerges from the Black Hills and is fed by cold water release from the hypolimnion of Pactola Reservoir. Since 2002, Dg blooms have been observed in certain stretches of the creek downstream of the reservoir. In this creek, the bloom is limited to stable low flow regimes covered with cobble and boulder type bed material. In many streams, most of the hot spots of the Dg algal bloom are mostly confined to downstream of regulated flow areas[1]. For example, in Rapid Creek, regardless of the homogeneity of the stream water chemistry (Fig. 2a and Table 1), Dg blooms are limited to low flow regimes located below Pactola Reservoir (Fig. 2b, c). Our result reveals that the relationship between the stream flow and biomass is hyperbolic and the maximum biomass (ash-free dry weight and chlorophyll-a) was measured under fairly low flow regimes (Fig. 2c). However, the biomass distribution was related to the residence time of the water parcel (Fig. 2c) and there is a strong positive relationship between water residence time and biomass (r2 = 0.78, p = 0.02). This can affect ion diffusion to and from water and biological matrix facilitating the mat development. Methods Results Site Description: To investigate the differential distribution of this algal bloom in Rapid Creek, we conducted measurements of stream flow, biomass, creek water and Dg mat tissue chemistry at 7 sites located below Pactola Reservoir (shown in Fig. 1) from 2008-2009 on a monthly basis from early spring to late fall, except during the winter, as the stream freezes during this time of the year. The reach of Rapid Creek from Pactola Reservoir to Rapid City has an average gradient of 48 ft/mi and is characteristic of a mountain stream with sand, gravel, and cobble bottom[5]. The stream crosses a series of predominantly sandstone and limestone which contains calcite, dolomite, and anhydrite[6].The creek is mainly characterized as hard (180 mg/L hardness as CaCO3[7], and alkaline pH (Fig. 2a). Conclusion The Dg tissue stoichiometry is fairly identical across the stations and seasons across flow regimes (Fig. 2d, e, and f). Since mat tissue chemistry is similar spatio-temporally, it is likely that the differential distribution of the Dg biomass bloom is influenced by hydrodynamics. Mat development itself alters water flow, creating low flow conditions around it, resulting in minimal surface water throughflow in the mat. This has direct impact on the microbial processes, chemistry of interstitial water in the mats, and redox stratification (Fig. 2g). While stream hydraulics is very important for mat stability (for example high flows can cause bed movement resulting in scouring of established Dg mats ), our results show that local hydrodynamic parameters, such as water residence time and hydrodynamic stablity, may be important predictors of Dg bloom distribution within a stream. c b) e) T28 Track c) f) T28 Track Table 1 g Figure 1. Map of the study site locations References 1. Elwell, L., and S. Spaulding, White paper. (January, 2007). 2. M. Miller et al., Hydrobiologia, 630, 207-218 (2009). 3. Bothwell, M.L. et.al., Fisheries, 34, 382-388 (2009). 4. Sundareshwar et al., 2010,(in press). 5. Williamson, J.E. et al., US Geological Survey, Water Resources Investigations Report 96-4276 (1996). 6. Peter, K.D., US Geological Survey, Water Resources Investigations Report 85-4022 (1995). 7. Freeman, J.E, and S.C.Komar, US Geological Survey Open File Report 91-249 (1991). Water Chemistry and Dg mat tissue analysis: Replicates of grab water samples were collected every month from 2008-2009. Creek water chemistry analysis was conducted using the Lachat automated nutrient analyzer by applying wet chemistry techniques. We measured silica, nitrate/nitrite, total nitrogen (TN), soluble reactive phosphorus (SRP) , total phosphorus (TP), and sulfate and sulfide(S2- ). Likewise, Dg mat total carbon (C) and Nitrogen (N) was conducted on CHN gas chromatography and total P was assayed using the wet digestion technique as described in Sundareshwar et al. (in press)[4]. The availability of major nutrients such as nitrogen and phosphorus is very low in this creek water (see Table 1). However, Dg tissue P is about 80% higher than creek water. Figure 2: Trends in (a) pH and temperature, (b) chlorophyll-a, (c) relationship between biomass (ash-free dry weight), water residence time, and stream flow, (d) TP in Dg mats, (e) seasonal changes in C, N & P content in Dg mats, along 8 stations on Rapid Creek, (f) N:P ratio in Dg mat versus Creek water, (g) Redox gradient in Dg mats incubated in laboratory. Table 1: Chemical characteristics of Creek water from Rapid Creek, SD and algal mats from Dg blooms in Rapid Creek. ND = No data. Acknowledgements This work was supported by funding from NSF DEB-0816593. We also wish to acknowledge Connie Crandall for her help in preparing this poster.