Download
1 / 32
320 likes | 485 Views
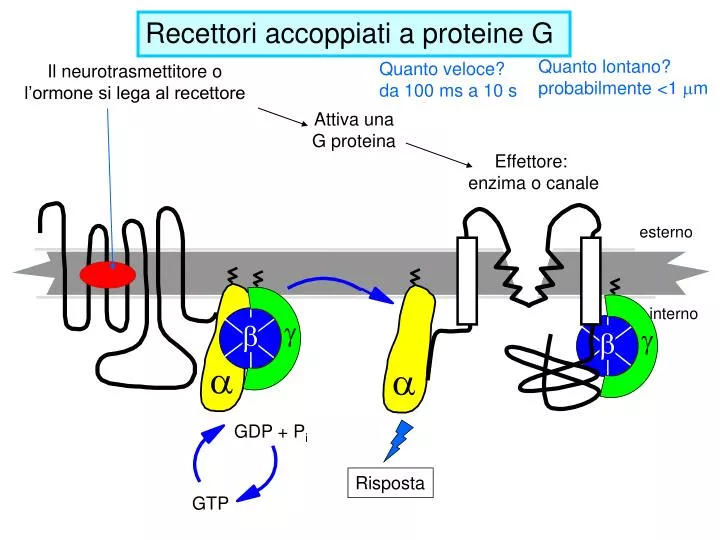
Quanto lontano? probabilmente <1 m m. Quanto veloce? da 100 ms a 10 s. Il neurotrasmettitore o l’ormone si lega al recettore. Attiva una G proteina. Effettore: enzima o canale. esterno. interno. a. a. g. g. GDP + P i. b. b. Risposta. GTP. Recettori accoppiati a proteine G.

E N D
Quanto lontano? probabilmente <1 mm Quanto veloce? da 100 ms a 10 s Il neurotrasmettitore o l’ormone si lega al recettore Attiva una G proteina Effettore: enzima o canale esterno interno a a g g GDP + Pi b b Risposta GTP Recettori accoppiati a proteine G
The alpha subunit of the heterotrimeric G protein is shown as a ribbon; the guanine nucleotide is spacefilled. P-alpha, P-beta, and P-gamma indicate the three phosphoryl groups in the GTP structure. As with most nucleoside triphosphates, there is a magnesium ion associated with GTP. The "ras-like" domain contains the catalytic residues that promote GTP hydrolysis.
The nucleotide binding site in each GTP-binding switch protein consists of loops that extend out from a b-sheet, usually 6-stranded. Three switch domainshave been identified, that change position when GTP substitutes for GDP on Ga. These include residues adjacent to the terminal phosphates of GTP and/or the Mg++ associated with them.
The bsubunit of the heterotrimeric G Protein has a b-propeller structure, formed from multiple repeats of a sequence called the WD-repeat. The b-propeller provides a stable structural support for residues that bind Ga.
The alpha subunit is usually modified by a fatty acyl lipid anchor. The gamma subunit is usually modified by an isoprenoid lipid anchor. Both lipid anchors (zig-zag lines) permit lateral diffusion, protein-lipid, and protein-protein interactions.
Outline the cyclic-AMP and phosphoinositide signal transduction cascades; differentiate between activation and inhibition of effector proteins by G protein subunits.
Turn off of the signal (when AC is activated): 1. Ga hydrolyzes GTP to GDP + Pi. (GTPase). The presence of GDP on Ga causes it to rebind to the inhibitory bgcomplex. Adenylate Cyclase is no longer activated. 2. Phosphodiesterasecatalyzes hydrolysis of cAMPAMP.
Turn off of the signal (cont.): 3. Hormone receptor desensitization occurs. This process varies with the hormone. • Some receptors are phosphorylated via G-protein-coupled receptor kinases (GRK). • The phosphorylated receptor may then bind to a protein arrestin that blocks receptor-G-protein activation and promotes removal of the receptor from the membrane by clathrin-mediated endocytosis. 4. Protein Phosphatase catalyzes removal by hydrolysis of phosphates that were attached to proteins via Protein Kinase A.
The Gs-alpha and Gi-alpha subunits both interact with adenylyl cyclase isoforms. Their actions, however, are opposite: Gs stimulates and Gi inhibits the synthesis of cyclic-AMP from ATP. The actions of these two alpha subunits may be differentiated in the laboratory by certain bacterial toxins.
Cholera toxin catalyzes covalent modification of Gsa. ADP-ribose is transferred from NAD+ to an arginine residue at the GTPase active site of Gsa. This ADP-ribosylation prevents Gsafrom hydrolyzing GTP. Thus Gsa becomes permanently activated. Pertussis toxin (whooping cough disease) catalyzes ADP-ribosylation at a cysteine residue of Gia, making the inhibitory Ga incapable of exchanging GDP for GTP. Thus the inhibitory pathway is blocked. ADP-ribosylation is a general mechanism by which activity of many proteins is regulated, in eukaryotes (including mammals) as well as in prokaryotes.
G proteins (guanine nucleotide (GTP) -binding proteins) G proteins, once activated, will cause the activation of several intracellulareffectors: adenyl cyclase, cGMP phosphodiesterase, phospholipase C, phospholipase A2, and calcium or potassium channels. lGi proteins (adenylate cyclase-inhibiting) - linked to a2-adrenergic receptor mGi1 protein mGi2 protein mGi3 protein lGo protein (Calcium or potassium channels modulators) lGq protein (Phospholipase C activator) linked to a1-adrenergic receptor lGs proteins (adenylate cyclase-stimulating) - linked to b-adrenergic receptor
Small GTP-binding proteins include (roles indicated): • initiation & elongation factors (protein synthesis). • Ras (growth factor signal cascades). • Rab (vesicle targeting and fusion). • Ran (transport of proteins into & out of the nucleus). • Rho (regulation of actin cytoskeleton) • All GTP-binding proteins differ in conformation depending on whether GDP or GTP is present at their nucleotide binding site. • Generally, GTP binding induces the active state.
Most GTP-binding proteins depend on helper proteins: GAPs, GTPase Activating Proteins, promote GTP hydrolysis. A GAP may provide an essential active site residue, and/or promote a conformation that favors catalysis. Ga of a heterotrimeric G protein has innate capability for GTP hydrolysis. However,RGS proteins, which are negative regulators of G protein signaling, function as GAPs to stimulate GTP hydrolysis by Ga.
GEFs, Guanine Nucleotide Exchange Factors, promote GDP/GTP exchange. The activated receptor (GPCR) serves as GEF for a heterotrimeric G protein.
The regulation of G protein signalling Left panel RGS proteins bind to G, stimulate GTP hydrolysis, and thereby reverse G protein activation. Right panel, the roles of a receptor, G, and an RGS are completely analogous to the GDSs, GDIs, and GAPs that regulate small monomeric G proteins like Ras.
G protein Mutations Causing Disease G protein alpha subunit in its GTP-bound form, highlighting amino acids changed by point mutations that cause human endocrine diseases. Mutational replacements of red residues impair GTP hydrolysis; these sites are mutated in growth hormone secreting tumors of the pituitary. Replacement of either cyan residue produces an inactive G protein alpha subunit, causing pseudohypoparathyroidism. Bound nucleotide is light green.

