Download

1 / 21
210 likes | 358 Views
Antibody domain exchange is an immunological solution to carbohydrate cluster recognition. Daniel Calarese … Ian A. Wilson (16 authors from Scripps, Oxford, FSU, U Ag Vienna) Science June 27, 2003 (300:2065). Burton, Wilson, and Calarese. What is “domain exchange”?. Paratope.

E N D
Antibody domain exchange is an immunological solution to carbohydrate cluster recognition. Daniel Calarese … Ian A. Wilson (16 authors from Scripps, Oxford, FSU, U Ag Vienna) Science June 27, 2003 (300:2065) Burton, Wilson, and Calarese slide by eric martz, sept 2003
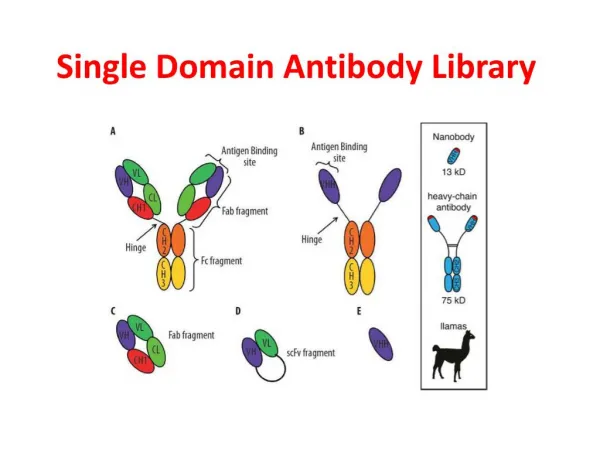
What is “domain exchange”? Paratope • 2 Ordinary Fabs • VH domains exchanged Forming a Fab dimer with two ordinary VH –VL paratopes + two new VH –VH paratopes Variable Domains Constant Domains H L slide by eric martz, sept 2003
Two Conven- tional Fab’s 2G12 Fab’s “Toober” models slide by eric martz, sept 2003
What is 2G12? • Human monoclonal IgG1 from patient B cells. • Neutralizes unusually broad range of known HIV strains (25-50%). • Binds to dense cluster of carbohydrate epitopes on the “silent” face of gp120. • Unusually high affinity among anti-carbohydrate antibodies. • Antibodies to the 2G12 epitope are unusual in sera from HIV+ patients (by competition). slide by eric martz, sept 2003
The “Silent” face of gp120 • Is an unusual epitope among HIV+ patient antibodies. • Potential protein epitopes beneath the carbohydrate are shielded. • Carbohydrate is made by host (not virus enzymes) hence may be “self” (though the dense cluster of oligomannose residues has not been described on mammalian glycoproteins). • Has multiple glycoforms diluting any one epitope. • Antibodies to carbohydrate epitopes usually have low affinities (Kd micromolar; but 2G12 Kd is nanomolar). slide by eric martz, sept 2003
Why is the structure of 2G12 important? • Paradigm shift: functional framework changes via somatic hypermutation. • Paradigm shift: VH-VH paratope. • Exception to the rule that Fab:antigen interactions are monovalent. • Domain-swapped dimer is unprecedented among >250 published Fab structures. • Explains high affinity and ability to bind closely-spaced epitopes. slide by eric martz, sept 2003
Protein Crystallography • Accounts for 85% of published structures. • Has an overall success rate of ~3%. • <1% of the human proteome has crystallographically known structure. slide by eric martz, sept 2003
Methods • Express full IgG1 in CHO cells. • Papain Fab (completeness confirmed by SDS-PAGE). • Purif. Prot. A/G columns, 30 mg/ml. • 1 μl (ligand 5 to 1 molar ratio) + 1 μl cryst. sol’n (ammonium/sulfate/phosphate, PEG, imidazole malate, pH 6/7), sitting drop crystallization. • Diffract Stanford Synchrotron cryopres. in Liq. N2 • Phase solved by molecular replacement with best-fitting existing Fab structure (1fvd). • Model of known amino acid sequence built into electron density map and refined (with restraints) for best fit. slide by eric martz, sept 2003
Crystallographic Results * ~6,600 atoms slide by eric martz, sept 2003
Framework Somatic MutationsFacilitate Domain Exchange • Weaken VH-VL • Alter H Chain Elbow Conserved ball and socket is not mutated but is dislocated. Mutated residues add stabilization. • Strengthen VH-VH’ See details in Protein Explorer slide by eric martz, sept 2003
Is this bizarre Fab dimer for real?It is not observed in >250 published Fab crystal structures. Fab: about 80% dimer + 20% monomer in solution by • Sedimentation equilibrium analytical ultracentrifugation. • Gel filtration. Intact IgG1 has compact conformation by • Velocity sedimentation coefficient. • Negative staining electron microscopy. • With and without gp140 ligand Apparently the dimer is for real. slide by eric martz, sept 2003
Fig. 2c: Electron microscopy of intact 2G12 IgG1 shows a compact form (neither Y nor T) with or without bound antigen. slide by eric martz, sept 2003
Mutants of 2G12reducing gp120 binding affinity ~100-1,000 fold • Primary combining site (VH-VL) 5/11 • Secondary binding site (VH-VH) 4/5 • Domain exchange-facilitating residues • VH-VH 4/4 • Elbow 3/3 slide by eric martz, sept 2003
Function of HIV oligomannose • DC-SIGN is a human lectin. (Dendritic Cell-Specific ICAM-3 Grabbing Nonintegrin) • Facilitates infection of CD4+ cells by binding HIV carbohydrate. • Speculation: HIV-1 evolved oligomannose to utilize DC-SIGN. • 2G12 exploits this “Achilles heel” of HIV-1. slide by eric martz, sept 2003
Conclusions • 2G12 forms a novel domain-swapped dimer (in 3 crystals and in solution) in both Fab and intact IgG1. • A novel form of paratope occurs at the VH-VH’ interface. • The novel conformation is enabled by somatic mutations to framework residues. • The resulting 4-paratope array accommodates closely spaced carbohydrate epitopes (35 Å, vs. 50-140 Å for Y or T) achieving nanomolar avidity. An epitope on gp120 is predicted. • Vaccines mimicking oligomannose cluster may elicit protective Abs. • 2IG2: scaffold for designing Abs to other epitope clusters? slide by eric martz, sept 2003
Alternative Antibody Lifestyles Naturally Occurring • Camel H chain dimer (no L; “VHH”) • (Muyldermans et al., TiBS 26:230, 2001) • VH domain-swapped dimer • (Calarese et al., Science 300:2065, 2003) Engineered • Fv (VH-VL with no C domains) • Fv domain-swapped “triabody” (trimer ring) • (Pei et al. PNAS 95:9637, 1997) slide by eric martz, sept 2003
Some High-Impact Crystal Structures • Myoglobin (1960): first example of protein structure. • Lysozyme (1965): first enzyme. • Fab (1973): first immunoglobulin fold (shared in unrelated proteins, first SOD). • tRNA (1975): first RNA structure. • MHC (1987): explained restriction of T cell recognition. • Ribosome (2000): peptidyl transferase is a ribozyme. • 2G12 (2003): unprecedented dimeric paratope with framework somatic mutations. slide by eric martz, sept 2003
First crystal structureswithout big surprises (less impact) • DNA double helix (1973-80): predicted correctly in 1953. • T cell antigen receptor (, 1996). • TCR (2001) • CD1 with phospholipid ligand (2002) slide by eric martz, sept 2003
Fig. 3D slide by eric martz, sept 2003
Fig. 3B slide by eric martz, sept 2003
Fig. 6: Model of 2G12 + gp120 slide by eric martz, sept 2003