Download
1 / 15
150 likes | 292 Views
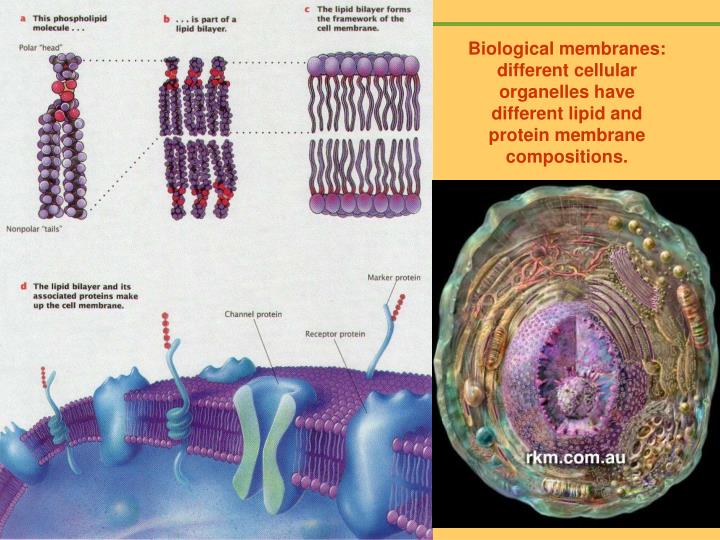
Biological membranes: different cellular organelles have different lipid and protein membrane compositions. Metabolic functions are often compartmentalized within membrane-bounded compartments in eukaryotic cells.

E N D
Biological membranes: different cellular organelles have different lipid and protein membrane compositions.
Metabolic functions are often compartmentalized within membrane-bounded compartments in eukaryotic cells. Although each cellular membrane has a unique set of proteins for its specialized functions, its basic building blocks are lipids. Lipids allow a snug fit of membrane-spanning proteins within the membrane, and ensure sufficient flexibility for vesicle budding and membrane fusion. Hydrophobic matching Many types of lipid are needed to determine precisely the physical properties of a given membrane in a cell. Moreover, cellular membranes contain many signalling lipids that transduce signals by interacting with specific proteins. Cells use both the structural and signalling properties of lipids to control membrane transport, thereby ensuring the unique molecular composition of each membrane-bounded compartment along the exocytic and endocytic pathways.
Lipids come in different shapes, which determines the self-assembled structures they prefer to form
Sorting: lipids sort proteins A membrane that contains mostly sphingomyelin,with or without cholesterol, is thicker than one composed of phosphatidylcholine and cholesterol, which is in turn thicker than a membrane of phosphatidylcholine alone. This implies that sphingolipid–cholesterol domains are thicker than the surrounding membrane. Cells probably use this feature to sort membrane proteins that are destined for the plasma membrane from Golgi proteins by the length of their transmembrane domains. For example, the transmembrane domains of plasma membrane proteins are 20 residues long, whereas those of Golgi proteins are only 15 residues long.
Protein-Induced Bilayer Deformations, and Lipid-Induced Protein Tilting
B Peptide-induced La to inverted phase transition A S. O. Nielsen et. al., Biophys. J. 87, 2107 (2004) in explanation of experimentally observed transition at 30:1 lipid:short-peptide concentration. C A. meniscus forms around peptides. B. water fills the meniscus regions. C. head groups rearrange to solvate the water pores.
Nature Reviews in Molecular Cell Biology, volume 2, page 504 (2001). We still have only sketchy information on the lipid composition of organellar membranes. In addition, our insight into the intracellular dynamics of lipids is limited. Finally, we know little about lipid–protein interactions at the molecular level, let alone, lipid– ipid interactions in complex mixtures.
We will introduce the new and important concept of protein/lipid sorting in membranes. It is suggested that in any membrane, the hydrophobic mismatch inherent to the protein and lipid composition may be released by a process of protein aggregation or, more interestingly, via a general mechanism of protein/lipid sorting. This concept of hydrophobic mismatch-dependent protein/lipid sorting is particularly attractive due to its inherent self- organizing character.
Hydrophobic mismatch induces formation of a meniscus • Maximal possible change in first shell lipid length is small and represent only a partial response to mismatch. (consistent with experiments, slope < 1) • Membrane thickens at intermediate range • The zero mismatch length can be obtained and compared to results obtained from free-energy calculations. S. O. Nielsen et. al., Biophys. J. 87, 2107 (2004) S. O. Nielsen et. al., Biophys. J. 88, 3822 (2005)
Free Energy Profile of Membrane Meniscus against Hydrophobic Mismatch Lipids do influence protein function—the hydrophobic matching hypothesis revisited (Biochimica et Biophysica Acta 2004, 1666, 205) Xibing He, S.O. Nielsen, et. al., manuscript in preparation: first time this effect has been quantified by computer simulation k = 650 kJ/mol
Lipid tilting J. Chem. Phys., 119, 7435 (2003) Lipid tilting in the vertical direction • Shorter tubes make the lipids adopt a more tilted and disorganized configuration (compared to bulk). • Longer tubes make lipids acquire a straighter configuration. • Straightening at intermediate distance due to thickening of membrane.
Lipid tilting Lipid tilting in the planar direction • For all peptide lengths the lipid’s head-to-tail vector points away from the peptide (due to rigid peptide, consistent with theory) with the shortest peptide giving the biggest tilting. • Region of negative correlation due to void resulting from positive meniscus (also seen by B. Smit)
Protein tilting Protein tilting from the bilayer normal Wide peptides are stable and do not tilt over the range of lengths studied. Tilting would deform the membrane too much (also reported in the DPD work of B. Smit) Narrow peptides tilt more as their length increases. The distribution also becomes bimodal.
Synthetic antimicrobials aryl amide oligomers Angew. Chem. Int. Ed., 43, 1158 (2004) Shai-Matsuzaki-Huang
How stable is the transmembrane orientation? Spontaneous insertion happens with the help of chaperone lipids. J. A. Killian has showed that TM peptides increase rate of flip-flop. C. F. Lopez, S. O. Nielsen, et. al., PNAS 101, 4431 (2004) This process can be quantified by computing the free energy of insertion.