Download
1 / 1
10 likes | 192 Views
Penes. Antenna 1. Antenna 2. Head width. Bodyshape. Pereopod 7. Pereopod 2. Uropod ekso. Uropod endo. App. masculina. 0.08. 2.8. T. macrura. 0.04. 2.4. T.smithi. 2.0. T. subequalum. 0.00. Penis length (size residuals). Appendix masculina length (mm). T. thermophilum. 1.6.

E N D
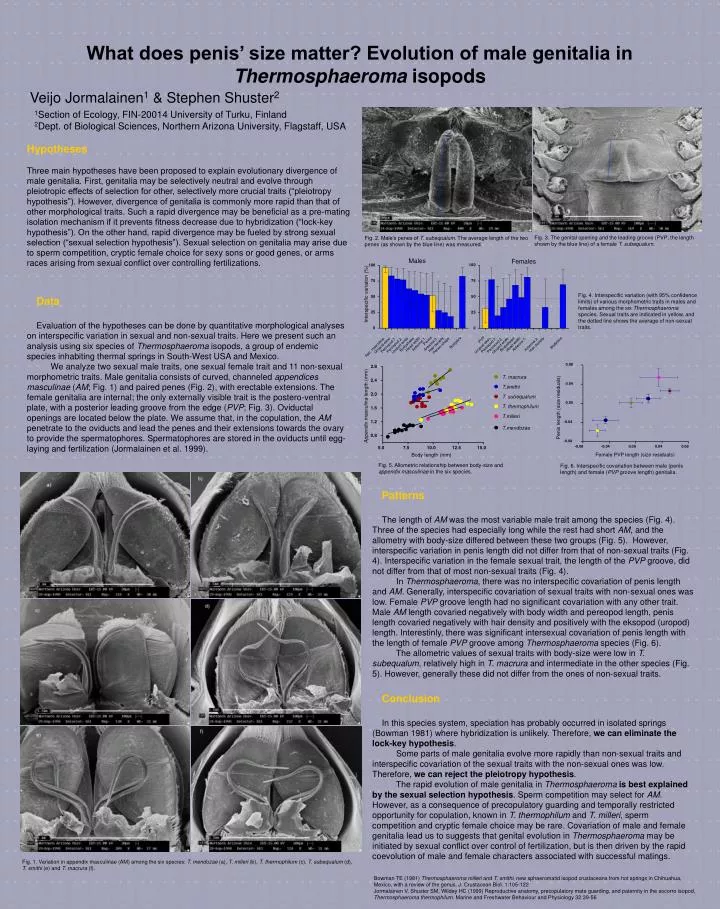
Penes Antenna 1. Antenna 2. Head width Bodyshape Pereopod 7. Pereopod 2. Uropod ekso Uropod endo App. masculina 0.08 2.8 T. macrura 0.04 2.4 T.smithi 2.0 T. subequalum 0.00 Penis length (size residuals) Appendix masculina length (mm) T. thermophilum 1.6 T.milleri -0.04 1.2 T.mendozae 0.8 -0.08 -0.08 -0.04 0.00 0.04 0.08 5.0 7.5 10.0 12.5 15.0 Female PVP length (size residuals) Body length (mm) What does penis’ size matter? Evolution of male genitalia in Thermosphaeroma isopods Veijo Jormalainen1 & Stephen Shuster2 1Section of Ecology, FIN-20014 University of Turku, Finland 2Dept. of Biological Sciences, Northern Arizona University, Flagstaff, USA Hypotheses Three main hypotheses have been proposed to explain evolutionary divergence of male genitalia. First, genitalia may be selectively neutral and evolve through pleiotropic effects of selection for other, selectively more crucial traits (“pleiotropy hypothesis”). However, divergence of genitalia is commonly more rapid than that of other morphological traits. Such a rapid divergence may be beneficial as a pre-mating isolation mechanism if it prevents fitness decrease due to hybridization (“lock-key hypothesis”). On the other hand, rapid divergence may be fueled by strong sexual selection (“sexual selection hypothesis”). Sexual selection on genitalia may arise due to sperm competition, cryptic female choice for sexy sons or good genes, or arms races arising from sexual conflict over controlling fertilizations. Fig. 3. The genital opening and the leading groove (PVP; the length shown by the blue line) of a female T. subequalum. Fig. 2. Male’s penes of T. subequalum. The average length of the two penes (as shown by the blue line) was measured. Males Females 100 100 75 75 Fig. 4. Interspecific variation (with 95% confidence limits) of various morphometric traits in males and females among the six Thermosphaeroma species. Sexual traits are indicated in yellow, and the dotted line shows the average of non-sexual traits. Interspecific variation (%) Data Evaluation of the hypotheses can be done by quantitative morphological analyses on interspecific variation in sexual and non-sexual traits. Here we present such an analysis using six speciesof Thermosphaeroma isopods, a group of endemic species inhabiting thermal springs in South-West USA and Mexico. We analyze two sexual male traits, one sexual female trait and 11 non-sexual morphometric traits. Male genitalia consists of curved, channeled appendices masculinae (AM; Fig. 1) and paired penes (Fig. 2), with erectable extensions. The female genitalia are internal; the only externally visible trait is the postero-ventral plate, with a posterior leading groove from the edge (PVP; Fig. 3). Oviductal openings are located below the plate. We assume that, in the copulation, the AM penetrate to the oviducts and lead the penes and their extensions towards the ovary to provide the spermatophores. Spermatophores are stored in the oviducts until egg-laying and fertilization (Jormalainen et al. 1999). 50 50 25 25 0 0 PVP Bodysize Bodysize Antenna 1. Antenna 2. Head width Bodyshape Hair density Hair density Pereopod 7. Pereopod 2. Uropod ekso Uropod endo Penes shape Fig. 5. Allometric relationship between body-size and appendix masculinae in the six species. Fig. 6. Interspecific covariation between male (penis length) and female (PVP groove length) genitalia. Patterns The length of AM was the most variable male trait among the species (Fig. 4). Three of the species had especially long while the rest had short AM, and the allometry with body-size differed between these two groups (Fig. 5). However, interspecific variation in penis length did not differ from that of non-sexual traits (Fig. 4). Interspecific variation in the female sexual trait, the length of the PVP groove, did not differ from that of most non-sexual traits (Fig. 4). In Thermosphaeroma, there was no interspecific covariation of penis length and AM. Generally, interspecific covariation of sexual traits with non-sexual ones was low. Female PVP groove length had no significant covariation with any other trait. Male AM length covaried negatively with body width and pereopod length, penis length covaried negatively with hair density and positively with the eksopod (uropod) length. Interestinly, there was significant intersexual covariation of penis length with the length of female PVP groove among Thermosphaeroma species (Fig. 6). The allometric values of sexual traits with body-size were low in T. subequalum, relatively high in T. macrura and intermediate in the other species (Fig. 5). However, generally these did not differ from the ones of non-sexual traits. Conclusion In this species system, speciation has probably occurred in isolated springs(Bowman 1981) where hybridization is unlikely. Therefore, we can eliminate the lock-key hypothesis. Some parts of male genitalia evolve more rapidly than non-sexual traits and interspecific covariation of the sexual traits with the non-sexual ones was low. Therefore, we can reject the pleiotropy hypothesis. The rapid evolution of male genitalia in Thermosphaeroma is best explained by thesexual selection hypothesis. Sperm competition may select for AM. However, as a consequence of precopulatory guarding and temporally restricted opportunity for copulation, known in T. thermophilum and T. milleri, sperm competition and cryptic female choice may be rare. Covariation of male and female genitalia lead us to suggests that genital evolution in Thermosphaeroma may be initiated by sexual conflict over control of fertilization, but is then driven by the rapid coevolution of male and female characters associated with successful matings. Fig. 1. Variation in appendix masculinae (AM) among the six species: T. mendozae (a), T. milleri (b), T. thermophilum (c), T. subequalum (d), T. smithi (e) and T. macrura (f). Bowman TE (1981) Thermosphaeroma milleri and T. smithi, new sphaeromatid isopod crustaceans from hot springs in Chihuahua, Mexico, with a review of the genus. J. Crustacean Biol. 1:105-122 Jormalainen V, Shuster SM, Wildey HC (1999) Reproductive anatomy, precopulatory mate guarding, and paternity in the socorro isopod, Thermosphaeroma thermophilum. Marine and Freshwater Behaviour and Physiology 32:39-56