Download

1 / 30
340 likes | 629 Views
Cell biology 2014 ( revised 21/1 -14), Note Lecture 2 handout. Ester bond. Lecture 2: . Membrane biology. Chapter 10 617-626 628-636. Chapter 11 651-664. Alberts et al. A lot of reading! Focus on principles and topics highlighted in the lecture synopsis . Chapter 12 695-699

E N D
Cell biology 2014 (revised 21/1 -14), Note Lecture 2 handout. Ester bond Lecture 2: Membrane biology Chapter 10 617-626 628-636 Chapter 11 651-664 Albertset al A lot of reading! Focus on principles and topics highlighted in the lecture synopsis Chapter 12 695-699 704-710 5th edition Cell Biologyinteractive media ”video” or ”interactive”
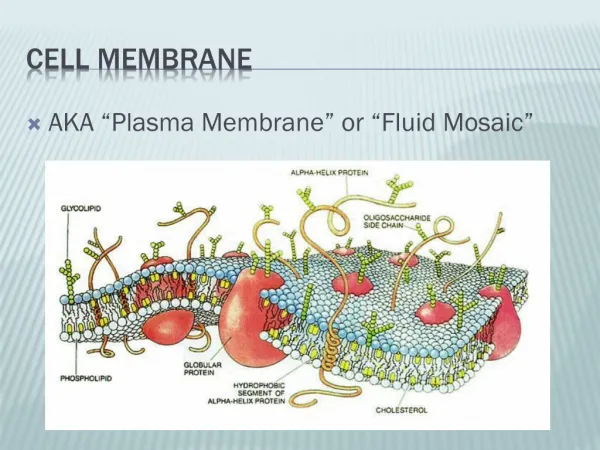
Membranes are primary built from phospholipids The major phospholipid: Phosphoglyceride Variable Lipid bilayer 5 -8 nm thick Hydrophilic head Phosphate Glycerol Biological membranes are lipid bilayers primary composed of amphipathicphospholipids Fatty acid Fatty acid Hydrophobic tails Glycerides(acylglycerols): esters formed from glycerol and fatty acids
Packing of amphipathic lipids in water - Wedge-shaped lipids form micelles in water H2O is a dipole Red: negative Blue: positive • Cylinder-shaped lipids form bilayers, followed by liposome • formation Amphipathiclipids will spontaneously form structures that eliminate the exposure of hydrophobic parts to water
Movement of individual lipids within the bilayer Rotational and lateral movement (frequent) Phospholipids can freely and rapidly (mm/s) diffuse within the monolayer Flip-flop (rare) Spontaneous movements between the two monolayers are rare The lipid bilayer is a two-dimensional fluid Similar viscosity as olive oil video 01.2 crawling_amoeba.mov; 13.5 phagocytosis .mov
van der Waals van der Waals van der Waals Fatty acid length affects membrane fluidity C=O C=O C=O C=O CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 Long fatty acid tails Short fatty acid tails Weak interactions High fluidity Strong interactions Low fluidity Long aliphatic carbon chains promote van der Waals interactions decreased membrane fluidity
Fatty acid saturation affects membrane fluidity An unsaturatedfattyacid has a kink Phospholipids containing only saturated fatty acids C=O C=O CH2 CH2 CH2 CH Phospholipids containing a unsaturated fatty acid CH CH2 CH2 CH2 CH2 CH2 Unsaturation's results in steric hindrance decreased van der Waals interactions increased membrane fluidity
Effect of lipid composition on membrane fluidity • Membrane thickness • Interactions between fatty acid chains - Membrane fluidity Shorter fatty acid chains and an increased degree of unsaturation make a thinner and more fluid lipid bilayer Anim. 09.1-laser_tweezer; Video 10.1- membrane_fluidity
Lipid rafts - clusters of strongly interacting lipids < 100 nm • The phospholipid sphingomyelin have long saturated fatty acid tails strong van der Waals interactions • Formation of a more static lipid environment Lipid rafts are micro-domains of phospholipids with low fluidity
Asymmetry of the plasma membrane Outer monolayer Inner monolayer (facing the cytosol) Phosphatidylcholine Phosphatidylethanolamine Phosphatidylserine Lipid raft former Sphingomyelin 50 40 30 20 10 0 10 20 30 40 50 Percentage of membrane lipids Extracellular space - - - - Phosphatidylinositol, important for cell signaling molecular_models 10.2-lipids.mov
1. 3. 2. 1. 3. 2. Different types of membrane proteins Peripheral Integral b-barrel Multi-pass a-helix Associated to Single-pass a-helix Mono-topic protein Lipid Integral membrane proteins are not tossed into the membrane randomly, but have a specific topology Integral protein Glycolipid
Dynamics of membrane proteins Rapid movement of proteins within the lipid bilayer Original fluid mosaic model (Singer& Nicolson 1972) Lipid raft Lipid micro-domain (Simons & Ikonen 1997) ~20 % of the plasma membrane
H R - O H N C C O H H + Membrane permeability of different molecules • Hydrophobic molecules O2 CO2 Benzene • Small unchargedpolar molecules H2O Ethanol • Large uncharged polar molecules Glucose H+ • Charged • molecules Na+ Cl- Ions Amino acids
Two types of transmembrane transport proteins Carrier Proteins Binds a “passenger” at one side of membrane and deliver it to the other side Channel proteins Creates a hydrophilic channel through the lipid bilayer that is selective for a particular solute From above
Ion channels • Most channel proteins are involved in ion transport over the • membrane and are therefore called ion channels • Ion channels are regulated and ion specific Closed Open Ion Ion A Ion B Ion Ion A Ion
Mechanisms behind membrane transport Facilitated specific diffusion Simple diffusion Active transport Concentration gradient Energy independent (down-hill) Energy dependent (up-hill)
P Different types of active membrane transport Transport of molecules against a concentration gradient requires energy. Cells uses two distinct strategies. Coupled transporters (symporters) ATP-driven pumps ATP ADP + “Up-hill” transport of molecule coupled to “down-hill” transport of molecule . The “down-hill” gradient depends on a ATP-driven pump “Up-hill” transport coupled directly to hydrolysis of ATP
P P P 3. 2. 1. 4. Example of active transport - Na+/K+ pump Na+ 145 mM K+ 5 mM Na+ Na+ Na+ Na+ Na+ Na+ Na+ 10 mM ADP ATP K+ 140 mM 1 cycle 10 milliseconds K+ K+ K+ K+ Anim. 11.2-carrier_proteins , Anim. 11.1-Na_K_pump
2. 1. 3. 2. 1. 3. Glucose Glucose Glucose Glucose Using concentration gradients of Na+ and K+ Active transport of Na+ and K+ creates concentration gradients The Na+ gradient provides the energy for “up-hill transport” Coupled transport of sucrose into the cytosol Na+ Na+ Na+ Na+ Na+ K+ Na+ K+ Na+ K+ K+ The ATP driving the Na+/K+ pump is the energy source for concentrating sugars and amino acids within cells
3. 2. 3. 1. 2. 1. Passive transport: facilitated “specific” diffusion of glucose to blood Glucose Glucose Glucose Glucose Glucose Glucose Glucose Example of trans-cellular transport by a symporter Na+ Na+ Intestinal lumen Na+ Na+ K+ K+ Blood vessels Na+ K+ K+ Na+/K+ pump establish Na+ gradient ATP Active transport: Na+ driven glucose symport (“cotransporter”) K+ Na+ Na+ Na+ Na+ Anim. 11.3-glucose_uptake
Compartments/organelles of eukaryotic cells Compartment Main function Cytosol Protein synthesis,metabolism Nucleus DNA & RNA synthesis Mitochondrion ATP production Endoplasmic Lipid synthesis, synthesis of proteins that reticulum (ER) enters the secretory pathway Golgi Sorting and packaging for delivery to cell surface or lysosome Lysosome Protein degradation
DNA replication Transcription mRNA, rRNA and tRNA Ribosome subunit assembly 1. 2. 3. 1. 2. 3. The nucleus – the instruction book of the cell Nuclear processes: Nuclear rRNA + proteins pore 3-10 mm
One reason for a nucleus in eukaryotes Prokaryote Eukaryote Transcription Transcription mRNA processing Translation Translation In eukaryotes mRNA has to be processed prior to initiation of translation, which requires spatial separation of transcription and translation (Note cloning of an ORF cDNA synthesis)
1. 2. 1. 2. Transport in and out of the nucleus Transcription mRNA tRNA Nuclear Nuclear rRNA pore pore DNA replication Protein synthesis in the cytosol
The nuclear pore complex (NPC) A typical cell contains 3000-4000 nuclear pore complexes 120 nm Annular subunit; the gatekeeper Outer nuclear membrane Inner nuclear membrane Proteins less than 60 kDa can diffuse ”freely” between cytosol and nucleus
Nuclear import of proteins (>60kD) Nuclear Localization Sequence (NLS) = sequence in a protein that mediates nuclear uptake N NLS C Could be localized anywhere in the protein N NLS C N NLS C Even distant apart in the primary structure of the protein N N L S C Which becomes adjacent in the folded protein N L S N C
NLS NLS NLS NLS 3. 4. 2. 1. 4. 2. 1. The process of facilitated nuclear protein import Nuclear import receptor (importin) Association of target protein and nuclear import receptor in the cytosol 3. Binding to the nuclear pore complex mediated by the nuclear import receptor ”Walking” through the gate-keepers of the pore Dissociation of target protein and nuclear import receptor inside the nucleus
GDP GDP Ran Ran 4. 3. 2. 1. The nuclear import cycle Cytosol Nucleus NLS Importin Importin NLS NLS GTP Importin Ran <60 kDa NLS +Pi Importin GTP Importin Ran GTP Importin Ran
GDP GDP Ran Ran The driving forces behind nuclear import NLS NLS Cytosol Nucleus Importin Importin NLS NLS <60 kDa Importin GDP GTP Energy cost! GTP Ran GTP GTP Importin Importin Ran Ran Video 02.3-brownian_motion.mov
<< GDP GTP Directionality in nuclear import – the Ran cycle Guanine-nucleotide Exchange Factor (GEF) G protein GTP GDP G protein GTPase Activating Protein (GAP) GDP GDP Ran Ran Pi Ran-GEF Ran-GAP GTP GTP Ran Ran Nucleus Cytosol
S Nuclear export Nuclear export of proteins is mediated by an intrinsic Nuclear Export Signal (NES). Proteins with NES include: Small protein that should not be nuclear NES Protein that shuttle between cytosol and nucleus NLS NES Video 12.2-nuclear_import.mov Export of mRNA is dependent on successful splicing N E S Proteins responsible for splicing Splicing; removal of introns from mRNA N E Spliced mRNA ready for nuclear export