Download
1 / 1
10 likes | 124 Views
Clustering & Phase Transitions on a Neutral Landscape A. SCOTT 1 , D. KING 1 , N. MARIĆ 2 , & S. BAHAR 1 1) Department of Physics & Astronomy and Center for Neurodynamics, University of Missouri at St Louis, St Louis, MO

E N D
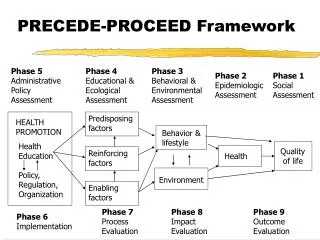
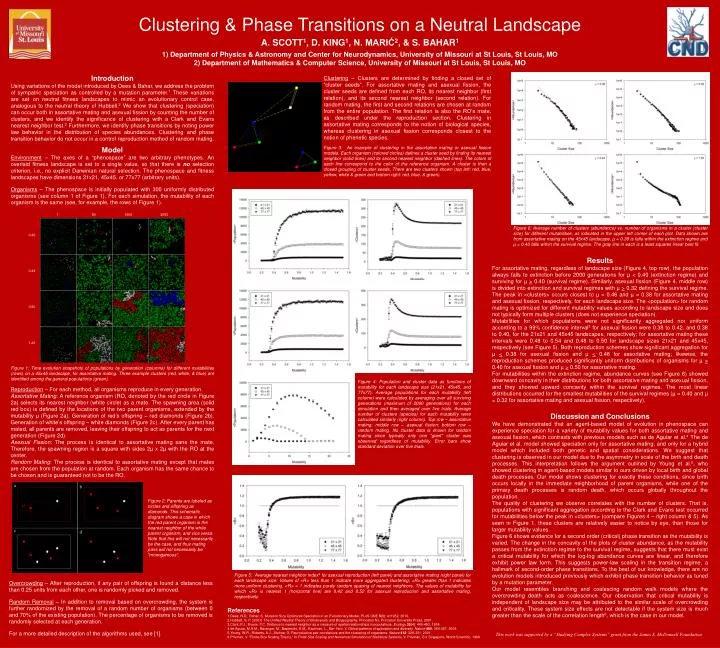
Clustering & Phase Transitions on a Neutral Landscape A. SCOTT1, D. KING1, N. MARIĆ2, & S. BAHAR1 1) Department of Physics & Astronomy and Center for Neurodynamics, University of Missouri at St Louis, St Louis, MO 2) Department of Mathematics & Computer Science, University of Missouri at St Louis, St Louis, MO 00.40 1 50 1000 2000 µ a b a b 00.44 Clustering – Clusters are determined by finding a closed set of “cluster seeds”. For assortative mating and asexual fission, the cluster seeds are defined from each RO, its nearest neighbor (first relation), and its second nearest neighbor (second relation). For random mating, the first and second relations are chosen at random from the entire population. The first relation is also the RO’s mate, as described under the reproduction section. Clustering in assortative mating corresponds to the notion of biological species, whereas clustering in asexual fission corresponds closest to the notion of phenetic species. Introduction Using variations of the model introduced by Dees & Bahar, we address the problem of sympatric speciation as controlled by a mutation parameter.1 These variations are set on neutral fitness landscapes to mimic an evolutionary control case, analogous to the neutral theory of Hubbell.2 We show that clustering (speciation) can occur both in assortative mating and asexual fission by counting the number of clusters, and we identify the significance of clustering with a Clark and Evans nearest neighbor test.3 Furthermore, we identify phase transitions by noting power law behavior in the distribution of species abundances. Clustering and phase transition behavior do not occur in a control reproduction method of random mating. 00.50 01.20 Figure 3: An example of clustering in the assortative mating or asexual fission models. Each organism (colored circles) defines a cluster seed by finding its nearest neighbor (solid lines) and its second nearest neighbor (dashed lines). The colors of each line correspond to the color of the reference organism. A cluster is then a closed grouping of cluster seeds. There are two clusters shown (top left: red, blue, yellow, white & green and bottom right: red, blue, & green). Model Environment –The axes of a “phenospace” are two arbitrary phenotypes. An overlaid fitness landscape is set to a single value, so that there is no selection criterion, i.e., no explicit Darwinian natural selection. The phenospace and fitness landscapes have dimensions 21x21, 45x45, or 77x77 (arbitrary units). Organisms–The phenospace is initially populated with 300 uniformly distributed organisms (see column 1 of Figure 1). For each simulation, the mutability of each organism is the same (see, for example, the rows of Figure 1). Figure 6: Average number of clusters (abundance) vs. number of organisms in a cluster (cluster size) for different mutabilities, as indicated in the upper left corner of each plot. Data shown are from assortative mating on the 45x45 landscape. µ = 0.38 is falls within the extinction regime and µ = 0.40 falls within the survival regime. The gray line in each is a least squares linear best fit. Results For assortative mating, regardless of landscape size (Figure 4, top row), the population always falls to extinction before 2000 generations for µ < 0.40 (extinction regime) and surviving for µ > 0.40 (survival regime). Similarly, asexual fission (Figure 4, middle row) is divided into extinction and survival regimes with µ > 0.32 defining the survival regime. The peak in <clusters> occurs closest to µ = 0.46 and µ = 0.38 for assortative mating and asexual fission, respectively, for each landscape size. The <population> for random mating is optimized for different mutability values according to landscape size and does not typically form multiple clusters (does not experience speciation). Mutabilities for which populations were not significantly aggregated nor uniform according to a 99% confidence interval3for asexual fission were 0.38 to 0.42, and 0.38 to 0.40, for the 21x21 and 45x45 landscapes, respectively; for assortative mating these intervals were 0.48 to 0.54 and 0.48 to 0.50 for landscape sizes 21x21 and 45x45, respectively (see Figure 5). Both reproduction schemes show significant aggregation for µ < 0.38 for asexual fission and µ < 0.48 for assortative mating; likewise, the reproduction schemes produced significantly uniform distributions of organisms for µ > 0.40 for asexual fission and µ > 0.50 for assortative mating. For mutabilities within the extinction regime, abundance curves (see Figure 6) showed downward concavity in their distributions for both assortative mating and asexual fission, and they showed upward concavity within the survival regimes. The most linear distributions occurred for the smallest mutabilities of the survival regimes (µ = 0.40 and µ = 0.32 for assortative mating and asexual fission, respectively). Figure 4: Population and cluster data as functions of mutability for each landscape size (21x21, 45x45, and 77x77). Average populations for each mutability (left column) were calculated by averaging over all surviving generations (maximum of 2000 generations) for each simulation and then averaged over five trials. Average number of clusters (species) for each mutability were calculated similarly (right column). Top row – assortative mating; middle row – asexual fission; bottom row – random mating. No cluster data is shown for random mating since typically only one “giant” cluster was observed regardless of mutability. Error bars show standard deviation over five trials. Figure 1: Time evolution snapshots of populations by generation (columns) for different mutabilities (rows) on a 45x45 landscape, for assortative mating. Three example clusters (red, white, & blue) are identified among the general populations (green). Reproduction – For each method, all organisms reproduce in every generation. Assortative Mating: A reference organism (RO, denoted by the red circle in Figure 2a) selects its nearest neighbor (white circle) as a mate. The spawning area (solid red box) is defined by the locations of the two parent organisms, extended by the mutability μ (Figure 2a). Generation of red’s offspring – red diamonds (Figure 2b). Generation of white’s offspring – white diamonds (Figure 2c). After every parent has mated, all parents are removed, leaving their offspring to act as parents for the next generation(Figure 2d). Asexual Fission: The process is identical to assortative mating sans the mate. Therefore, the spawning region is a square with sides 2µ x 2µ with the RO at the center. Random Mating: The process is identical to assortative mating except that mates are chosen from the population at random. Each organism has the same chance to be chosen and is guaranteed not to be the RO. Discussion and Conclusions We have demonstrated that an agent-based model of evolution in phenospace can experience speciation for a variety of mutability values for both assortative mating and asexual fission, which contrasts with previous models such as de Aguiar et al.4 The de Aguiar et al. model showed speciation only for assortative mating, and only for a hybrid model which included both genetic and spatial considerations. We suggest that clustering is observed in our model due to the asymmetry in scale of the birth and death processes. This interpretation follows the argument outlined by Young et al.5, who showed clustering in agent-based models similar to ours driven by local birth and global death processes. Our model shows clustering for exactly these conditions, since birth occurs locally in the immediate neighborhood of parent organisms, while one of the primary death processes is random death, which occurs globally throughout the population. The quality of clustering we observe correlates with the number of clusters. That is, populations with significant aggregation according to the Clark and Evans test occurred for mutabilities below the peak in <clusters> (compare Figures 4 – right column & 5). As seen in Figure 1, these clusters are relatively easier to notice by eye, than those for larger mutability values. Figure 6 shows evidence for a second order (critical) phase transition as the mutability is varied. The change in the concavity of the plots of cluster abundance, as the mutability passes from the extinction regime to the survival regime, suggests that there must exist a critical mutability for which the log-log abundance curves are linear, and therefore exhibit power law form. This suggests power-law scaling in the transition regime, a hallmark of second-order phase transitions. To the best of our knowledge, there are no evolution models introduced previously which exhibit phase transition behavior as tuned by a mutation parameter. Our model resembles branching and coalescing random walk models where the overcrowding death acts as coalescence. Our observation that critical mutability is independent of landscape size may be attributed to the similar scale of overcrowding and criticality. These system size effects are not detectable if the system size is much greater than the scale of the correlation length6, which is the case in our model. a b Figure 2: Parents are labeled as circles and offspring as diamonds. This schematic diagram shows a case in which the red parent organism is the nearest neighbor of the white parent organism, and vice versa. Note that this will not necessarily be the case, and thus mating pairs will not necessarily be "monogamous“. c d Figure 5: Average nearest neighbor index3for asexual reproduction (left panel) and assortative mating (right panel) for each landscape size. Values of <R> less than 1 indicate more aggregated clustering; <R> greater than 1 indicates more uniform clustering. <R> = 1 indicates purely random spacing of nearest neighbors. The values of mutability for which <R> is nearest 1 (horizontal line) are 0.42 and 0.52 for asexual reproduction and assortative mating, respectively. Overcrowding – After reproduction, if any pair of offspring is found a distance less than 0.25 units from each other, one is randomly picked and removed. Random Removal – In addition to removal based on overcrowding, the system is further randomized by the removal of a random number of organisms (between 0 and 70% of the existing population). The percentage of organisms to be removed is randomly selected at each generation. For a more detailed description of the algorithms used, see [1]. • References • Dees, N.D., Bahar, S. Mutation Size Optimizes Speciation in an Evolutionary Model. PLoS ONE 5(8): e11952, 2010. • Hubbell, S. P. (2001) The Unified Neutral Theory of Biodiversity and Biogeography. Princeton NJ, Princeton University Press, 2001. • Clark, P.J., Evans, F.C. Distance to nearest neighbor as a measure of spatial relationships in populations. Ecology35(4): 445-453, 1954. • de Aguiar, M.A.M., Baranger, M., Baptestini, E.M., Kaufman, L., Bar-Yam, Y. Global patterns of speciation and diversity. Nature460: 384-387, 2009. • Young, W.R., Roberts, A.J., Stuhne, G. Reproductive pair correlations and the clustering of organisms. Nature412: 328-331, 2001. • Privman, V. "Finite-Size Scaling Theory." In Finite Size Scaling and Numerical Simulation of Statistical Systems, V. Privman, Ed. Singapore, World Scientific, 1990. This work was supported by a “Studying Complex Systems” grant from the James S. McDonnell Foundation.