Download

1 / 31
370 likes | 1.28k Views
Drosophila anterior-posterior axis formation during early embryogenesis. Genetics Unit Department of Biochemistry petros.ligoxygakis@bioch.ox.ac.uk. Developmental biology: Drosophila segmentation and repeated units. * egg: generate the system. * larva: eat and grow.

E N D
Drosophila anterior-posterior axis formation during early embryogenesis Genetics Unit Department of Biochemistry petros.ligoxygakis@bioch.ox.ac.uk
Developmental biology: Drosophila segmentation and repeated units * egg: generate the system * larva: eat and grow * pupa: structures in larvae grow out to form adult fly: metamorphosis (Drosophila is a holometabolous insect) 1
The origins of Anterior-Posterior polarity • The anterior-posterior polarity of the embryo, larva and adult has it’s origin in the anterior posterior polarity of the egg. The maternal effect genes expressed in the mothers ovaries produce mRNAs that are placed in different regions of the egg. These messages encode transcriptional and translational regulatory proteins that diffuse through the syncytial blastoderm and activate or repress the expression of certain zygotic genes
Axes in the Drosophila embryo are specified in the egg • Cooperation between nurse cells, which make the maternal mRNAs and follicle cells that signal to the oocyte and reorganise its cytoskeleton, is essential for localisation of bicoid and nanos
EGF signalling between the oocyte nucleus and follicle cells
At the end of signalling posterior and dorsal follicle cells have obtained a different identity
Different cytoskeletal systems in the same cell Intermediate filaments (vimentin) Microtubules (tubulin) Microfilaments (actin) The cytoskeleton performs several tasks that are importa. nt for the formation of assymetries: control of the location of the mitotic cleavage plane within the cell, control of cell shape and directed transport of molecules and organelles within the cell. All of these tasks depend on the fact that these rods are polar structures exhibiting directionality. That is one end of the cytoskeletal rod can be chemically distinguished from the other
Distribution of tubulin in animal cells Microtubules radiate out from a microtubule organising centre. The negative (minus) ends of the microtubules are in the centre and the positive (plus) ends are at the periphery of the cell.
Motors and cargoes Different motors are used for travelling to different directions carrying different cargoes. For example dynein can travel in the opposite direction than kinesin. What’s the value of having multiple independent trafficking systems? A part of the answer is division of labour.
Nuclear divisions in the syncytial embryo • It is important to note that in the early stages in Drosophila embryonic development there are nuclei divisions inside the embryonic cytoplasm (the syncytium). This is important in order to understand how in these stages transcription factors are used to drive the gene regulation cascade, which will lead to the final patterning of the embryo after 24h D P A V
Drosophila embryonic development in real time • Most insect eggs undergo superficial cleavage, where in a large number of mass of centrally located yolk confines cleavage to the cytoplasmic rim of the egg. During these early stages of division there are no cells formed but the embryo is a syncytial blastoderm meaning that the egg is a large cell containing within a common cytoplasm the dividing nuclei. Following division cycle 13 the oocyte plasma membrane folds inward between the nuclei and cellularisation begins. D A P V
Breaking symmetry: the way to establish positional information • A) Create a population of developmentally identical cells • B) Create an asymmetry within that population • C) Exploit the asymmetry to set up a chemical concentration gradient within the population of cells • D) Different cells will mount different responses to local concentrations of these gradients • E) Different responses will create different states of developmental potential
Pattern formation in Drosophila • A) the cytoskeleton imposes an asymmetry on the egg. • B) Gradients of soluble proteins are laid down along the A-P and the D-V axis, stimulating gene action and • C) leading to broad regions of gene expression.
The Heidelberg screen • Christiane Nüsslein-Volhard, Eric Wieschaus and colleagues performed extensive mutational screens, essentially saturating the Drosophila genome for mutations that alter the A-P and D-V patterns of the larva exoskeleton with the assumption that such alterations would reflect changes in the basic body plan.
Mutational analysis of Drosophila early embryogenesis • Two broad classes of genes affecting the body pattern: zygotic genes, which are expressed exclusively in the zygote being part of its DNA and maternal genes, which are contributed by the mother and the phenotype of the offspring depends on the phenotype of the mother.
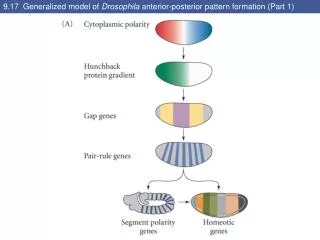
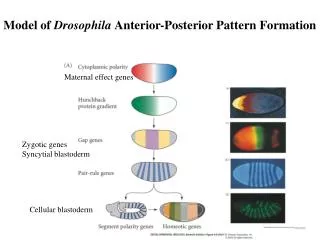
Concentration gradients of BCD and HB-M establish A-P axis • Positional information along the A-P axis of the syncitial embryo is initially established through the creation of concentration gradients of two transcription factors: Bicoid (BCD) and Hunchback (HB-M). These are products of two maternal effect genes their mRNAs provided by the mother and stored in the embryo until translation initiates. These factors interact to generate different patterns of gene expression along the axis.
Localisation of A-P determinants in the early embryo In situ hybridisation (RNA) Antibody detection (protein)
Effect of replacement of the 3’ UTR of the nos mRNA with the 3’ UTR of bcd mRNA • The nos-bcd transgene is able to localise at the anterior pole and as a consequence NOS protein will inhibit translation of the hb and bcd mRNAs.
MESSAGE: Positional information in Drosophila embryonic A-P axis is generated by protein gradients. The gradients ultimately depend on diffusion of newly translated protein from a localised source of mRNAs anchored by their 3’ UTR to ends of cytoskeleton filaments
Studying the BCD gradient I • In embryos derived from bcd mothers the anterior (head and thorax) part of the body is missing (a). This can be restored by injection of either anterior cytoplasm (b) or the actual wild type mRNA (c).
Studying the BCD gradient II • The amount of BCD protein can be changed by varying copies of bcd+ in the mother. This changes the position of the gradient to more posterior regions.
SUMMARY Nurse cells surrounding the oocyte in the ovarian follicle provide it with large amounts of mRNAs and proteins, some of which become localised in particular sites. The oocyte produces a local signal, which induces follicle cells at one end to become posterior follicle cells. The posterior follicle cells cause a re-organisation of the oocyte cytoskeleton that localises bicoid and hunchback mRNA to the anterior end and other mRNAs such as oskar and nanos to the posterior end of the oocyte. Following fertilisation, development starts and these mRNAs are translated. Subsequently, gradients of the BCD and HB proteins define the anterior nuclei-the embryo is still a syncytial blastoderm, while inhibition of translation of their mRNAs by Nanos define the posterior cells. Nuclei in between receive a variable amount of BCD and HB resulting in differential activation or repression of target genes and finally in different developmental cell fates.
READING LIST Wolpert et al, Principles of Development St Johnston D and Nusslein-Volhard C (1992) The origin of pattern and polarity in the Drosophila embryo Cell 68, 201-219 St Johnston D (2005) Moving messages: the intracellular localisation of mRNAs Nat Rev Mol Cell Biol 6, 363-375. Further reading: Zimyanin et al, 2008 In vivo imaging of oskar mRNA transport reveals the mechanism of posterior localisation Cell 134, 843-853