Download
1 / 46
460 likes | 645 Views
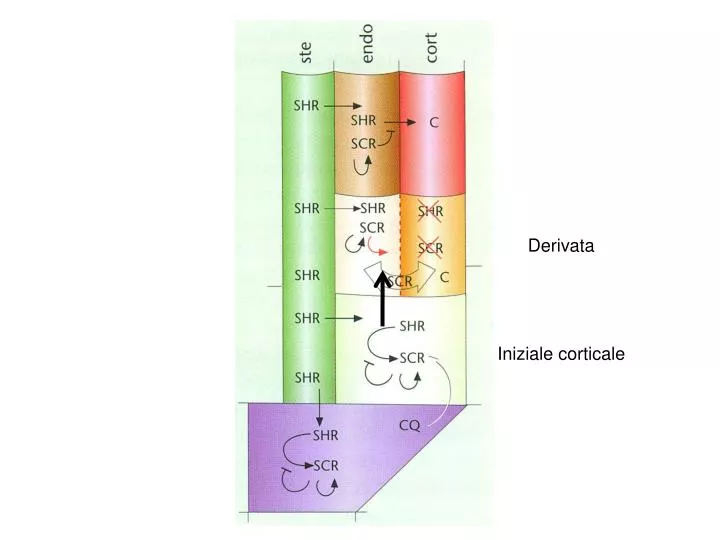
Derivata Iniziale corticale. Funzione dei geni SHORT ROOT e SCARECROW Appartengono alla famiglia GRAS (GAI, RGA) di fattori di trascrizione. G IBBERELLIN-INSENSITIVE (GAI) R EPRESSOR OF ga1-3 (RGA) S CR. Struttura GAI e RGA. RGA ( rga) isolato come soppressore del fenotipo ga1.

E N D
Derivata Iniziale corticale
Funzione dei geni SHORT ROOT e SCARECROW Appartengono alla famiglia GRAS (GAI, RGA) di fattori di trascrizione GIBBERELLIN-INSENSITIVE (GAI) REPRESSOR OF ga1-3 (RGA) SCR
RGA (rga) isolato come soppressore del fenotipo ga1 ga1 è un mutante della via di biosintesi delle GA (ha livelli ridotti di GA) e quindi è nano
RGA ha un’identità dell’82% con GAI Tuttavia la mutazione gai determina un fenotipo nano
Le mutazioni gai e rga sono in domain diversi delle proteine gai-1: delezione del dominio DELLA (17 aa) rga: mutazione nel dominio repressore GRAS
RGA e GAI sono dei regolatori negativi della risposta alle gibberelline Le GA inducono la degradazione di RGA e GAI
DIFFERENZIAMENTO DI XILEMA E FLOEMA PROCAMBIO: genera il protoxilema e il protofloema e successsivamente il metaxilema ed il metafloema. Il differenziamento è centripeto. Radice di arabidopsis in struttura primaria In arabidopsis il differenziamento in metaxilema arriva fino al centro del cilindro vascolare (manca il midollo)
WOODEN LEG (WOL) Mutanti wol hanno un sistema vascolare costituito da solo Xilema e un numero ridotto di file cellulari
- +
CKs control root vascular development In the root vascular cylinder, some cells in the procambium differentiate into protoxylem cells (yellow arrows), others into metaxylem (yellow arrowheads) and others into phloem (red arrowheads). The cell walls of protoxylem have a characteristic ringed structure not present in the metaxylem. From Mahonen, A.P., et al (2006). Cytokinin signaling and its inhibitor AHP6 regulate cell fate during vascular development. Science 311: 94-98.Reprinted with permission from AAAS.
CKs control root vascular development In the wooden leg (wol) mutant of AHK4 all the vascular cells differentiate to protoxylem. From Mahonen, A.P., et al (2006). Cytokinin signaling and its inhibitor AHP6 regulate cell fate during vascular development. Science 311: 94-98.Reprinted with permission from AAAS.
A triple mutant of type-B ARRs or ahks causes the same phenotype Protoxylem differentiation causes the root to stop growing. Argyros, R.D.,et al. (2008). Type B response regulators of Arabidopsis pay key roles in cytokinin signaling and plant development. Plant Cell 20: 2102-2116.
Overexpression of ARR22 causes a wol-like phenotype These results suggests that the wol mutation causes a severe inhibition of CK signaling. Kiba, T., Aoki, K., Sakakibara, H., and Mizuno, T. (2004). Arabidopsis Response Regulator, ARR22, ectopic expression of which results in phenotypes similar to the wol cytokinin-receptor mutant. Plant Cell Physiol. 2004 45: 1063-1077
CK prevents cells from differentiating into protoxylem CK Procambium Other Proto-xylem Adapted from Mahonen, A.P. et al (2006). Cytokinin signaling and its inhibitor AHP6 regulate cell fate during vascular development. Science 311: 94-98.
A loss-of-function ahp6 mutant partially rescues the wol mutant AHP6 is expressed in the procambium cells destined to form protoxylem wol/ ahp6 WT wol CK AHP6 Pro-cambium Proto-xylem AHP6 interferes with CK signaling to permit protoxylem development From Mahonen, A.P., et al (2006). Cytokinin signaling and its inhibitor AHP6 regulate cell fate during vascular development. Science 311: 94-98.Reprinted with permission from AAAS.
The wol phenotype is MORE severe than a loss-of-function mutation WT wol WT wolahk4 (LOF) The woodenleg (wol) mutant allele is more severe than a loss-of-function ahk4 mutation. Instead, it resembles a loss –of-function of all three CK receptors ahk4 ahk2,3,4 * * * * * * * * * * * * * * * * * * * * * *
Why is the wol mutation so severe? AHK2 H H H D D D D AHK3 ahk4 H Loss-of-function of the AHK4 receptor does not stop CK signaling. Why does the wol mutation stop CK signaling?
The wol mutation has a constitutive phosphatase activity AHK2 H H H D D D D AHK3 wol H A point mutation switches the mutant wol protein into a constitutive phosphatase that, like ARR22, removes phosphoryl groups from the phosphorelay system.
Studies of the wol mutant have revealed that: CK responses can be controlled temporally and spatially to specify developmental fates. Bidirectional transfer of phosphoryl groups and phosphatase activities add complexity (and additional controls) to the phosphorelay system.
DIFFERENZIAMENTO DI XILEMA E FLOEMA • Quattro aspetti principali: • Formazione continua del procambio durante lo sviluppo (polarità apice/base) • Generazione della asimmetria attraverso il procambio (polarità radiale) • Bilancio tra proliferazione cellulare e differenziamento dei due tipi cellulari • Acquisizione della identità xilematica o floematica in accordo alla asimmetria del procambio
Formazione continua del procambio durante lo sviluppo Tessuto più interno nel pattern radiale Evento primario Evidenze che il processo sia innescato dal trasporto polare dell’auxina (polarità apice/base)
Generazione della asimmetria nel procambio • strettamente associata alla generazione della polarità negli organi • (adaxiale/abaxiale nelle foglie e centrale /periferica nel fusto) “BATTAGLIA” tra fattori HD-ZIPIII e geni KANADI
La generazione del’’asimmetria nel procambio è regolata dalla interazione tra HD-ZIPIII e GENI KANADI
HD-ZIP III • REVOLUTA/INTERFASCICULAR FIBERLESS (REV) • PHABULOSA/ATHB14 (PHB) • PHAVOLUTA (ATHB9) • CORONA/ATB15 (CAN) • ATHB8
GENE PHANTASTICA di ANTHIRRINUM (fattore MYB) Nelle foglie mutazioni loss of function phan promuovono la conversione adaxiale/abaxiale I trascritti sono localizzati nel lato adaxiale
mutazione phan in Anthirrinum, l’ortologo in arabidopsis è Asimmetric Leaves 1 (AS1) determina la conversione adaxiale/abaxiale gene di identità adaxiale
In arabidopsis mutanti phan o tripli mutanti rev/phb/phv Sviluppano cotiledoni simmetrici radialmente con vascolatura abaxializzata
Mutazioni gain-of-function dei geni HD-ZIPIII producono organi laterali adaxializzati e sviluppano fasci vascolari in cui il floema è circondato da xylema
Le mutazioni sono situate in una sequenza target per miRNA165 o miRNA166 all’ interno di un dominio START (sterol/lipid binding domain) miR165 si accumula nel dominio abaxiale/floematico della foglia; pattern complementare a quello di PHB
Phabulosa; Phavoluta; Revoluta Homeodomain leucin zipper proteins HD-ZIPIII contenenti un dominio START che lega lipidi Fattori di trascrizione specifici per il lato adaxiale
La localizzazione nella regione adaxiale dei trascritti HD-ZIP è regolata da: Geni KANADI (KAN) espressi nella regione abaxiale (mutazioni recessive kan determinano adaxializzazione e espressione ectopica degli HD-ZIP)
miR165 e miR166 sono espressi nel lato abaxiale delle foglie e nel floema Pattern analogo a quello dei geni KANADI I geni KANADI controllano l’espressione di miR165/166 REPRESSIONE dell’espressione dei fattori HD-ZIPIII
KANADI HD-ZIP III (miRNA) HD-ZIP III
I mutanti kan1, kan2, kan 3 fenocopiano i mutanti gain-of-function HD-ZIPIII
proliferazione/differenziamento cellule floematiche e xilematiche I brassinosteroidi influenzano il differenziamento vascolare
BRL1, BRL3 In arabidopsis identificati due nuovi recettori dei brassinosteroidi omologhi a BRI1 (LRR-receptor Kinase) espressi nel tessuto vascolare L’analisi dei mutanti suggerisce che essi siano ridondanti e che inducano il differenziamento dello xilema e reprimano la proliferazione del floema Quali geni sono attivati dalla via di signaling dei brassinolidi?
In cellule del mesofillo di Zinnia elegans i BR inducono il differenziamento in trachee Il differenziamento coincide con l’espressione di omologhi dei geni HD-ZIPIII di arabidopsis (ATHB8, ATHB15, REV)
In arabidopsis l’overespressione di ATHB8 induce proliferazione del tessuto vascolare e differenziamento dello xilema HD-ZIP III : regolatori positivi della proliferazione e del differenziamento del tessuto vascolare
ACQUISIZIONE IDENTITA’ XILEMA/FLOEMA • ALTERED PHLOEM DEVELOPMENT (APL): fattore MYB • Espresso nel floema • In mutanti apl (letali)si hanno elementi xilematici al posto dei floematici; • espressione ectopica di APL nella stele sopprime l’espressione del protoxilema • e di parte del metaxilema ma il differenziamento delle cellule floematiche • è incompleto Necessario ma non sufficiente per il differenziamento in floema
XILOGENO: identificato in trachee immature di Zinnia come un fattore di secrezione che promuove il differenziamento di cellule vicine Piccola proteina con caratteristiche di arabinogalattanoproteina e lipid transfer protein (LPT) È localizzato in maniera polare nella parte apicale di tracheee immature Forse può interagire con steroli tramite il dominio delle LTP
