Download
1 / 12
120 likes | 317 Views
Cell cycle. Lesson 11. Cell cycle. The cell cycle is the cyclical process by which a cell that has just divided grows , replicates its DNA, and then divides again. In eukaryotes, it is generally considered to have four phases:

E N D
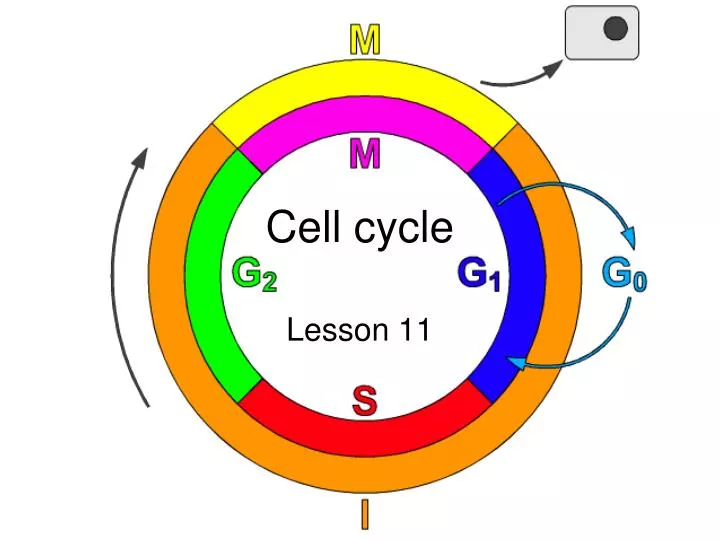
Cell cycle Lesson 11
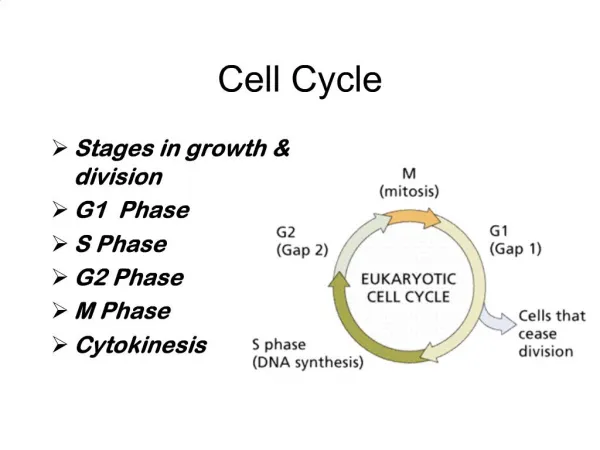
Cell cycle • The cell cycle is the cyclical process by which a cell that has just divided grows, replicates its DNA, and then divides again. In eukaryotes, it is generally considered to have four phases: • G1 - the first gap phase, during which the cell grows to a threshold size. The cell actively takes up materials from its environment to form new organelles and cytoplasm. • S - during which the cell replicates its DNA and MTOCs (microtubule organising centres: i.e. centrioles, centrosomes, etc.) • G2 - (collectively known as interphase) the second gap phase, during which the cell condenses its genome and MTOCs begin to form a spindle which will later separate the chromosomes. • M - mitosis (or meiosis), during which the genome is passive, whilst the cytoskeleton actively moves the chromosomes about. http://www.steve.gb.com/science/cell_cycle.html
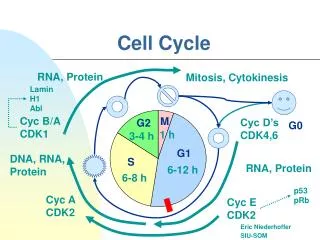
Phases of the cell cycle • M phase: The relatively brief M phase consists of nuclear division (karyokinesis) and cytoplasmic division (cytokinesis). In plants and algae, cytokinesis is accompanied by the formation of a new cell wall. • Interphase: After M phase, the daughter cells each begin interphase of a new cycle. Although the various stages of interphase are not usually morphologically distinguishable, each phase of the cell cycle has a distinct set of specialized biochemical processes that prepare the cell for initiation of cell division. • G1 phase: The first phase within interphase, from the end of the previous M phase till the beginning of DNA synthesis is called G1 (G indicating gap or growth). During this phase the biosynthetic activities of the cell, which had been considerably slowed down during M phase, resume at a high rate. This phase is marked by synthesis of various enzymes that are required in S phase, mainly those needed for DNA replication. Duration of G1 is highly variable, even among different cells of the same species. • S phase: The ensuing S phase starts when DNA synthesis commences; when it is complete, all of the chromosomes have been replicated, i.e., each chromosome has two (sister) chromatids. Thus, during this phase, the amount of DNA in the cell has effectively doubled, though the ploidy of the cell remains the same. Rates of RNA transcription and protein synthesis are very low during this phase. An exception to this is histone production, most of which occurs during the S phase. The duration of S phase is relatively constant among cells of the same species. • G2 phase: The cell then enters the G2 phase, which lasts until the cell enters mitosis. Again, significant protein synthesis occurs during this phase, mainly involving the production of microtubules, which are required during the process of mitosis. Inhibition of protein synthesis during G2 phase prevents the cell from undergoing mitosis. • G0 phase: The term "post-mitotic" is sometimes used to refer to both quiescent and senescent cells. Nonproliferative cells in multicellular eukaryotes generally enter the quiescent G0 state from G1 and may remain quiescent for long periods of time, possibly indefinitely (as is often the case for neurons). This is very common for cells that are fully differentiated. Cellular senescence is a state that occurs in response to DNA damage or degradation that would make a cell's progeny nonviable; it is often a biochemical alternative to the self-destruction of such a damaged cell by apoptosis. Some cell types in mature organisms, such as parenchymal cells of the liver and kidney, enter the G0 phase semi-permanently and can only be induced to begin dividing again under very specific circumstances; other types, such as epithelial cells, continue to divide throughout an organism's life. Schematic of the cell cycle. outer ring: I=Interphase, M=Mitosis; inner ring: M=Mitosis, G1=Gap 1, G2=Gap 2, S=Synthesis; not in ring: G0=Gap 0/Resting. The duration of mitosis in relation to the other phases has been exaggerated in this diagram.
Regulation of cell cycle • Regulation of the cell cycle involves steps crucial to the cell, including detecting and repairing genetic damage, and provision of various checks to prevent uncontrolled cell division. The molecular events that control the cell cycle are ordered and directional; that is, each process occurs in a sequential fashion and it is impossible to "reverse" the cycle. • Role of Cyclins and CDKs • Two key classes of regulatory molecules, cyclins and cyclin-dependent kinases (CDKs), determine a cell's progress through the cell cycle. Leland H. Hartwell, R. Timothy Hunt, and Paul M. Nurse won the 2001 Nobel Prize in Physiology or Medicine for their discovery of these central molecules. Many of the genes encoding cyclins and CDKs are conserved among all eukaryotes, but in general more complex organisms have more elaborate cell cycle control systems that incorporate more individual components. Many of the relevant genes were first identified by studying yeast, especially Saccharomyces cerevisiae; genetic nomenclature in yeast dubs many of these genes cdc (for "cell division cycle") followed by an identifying number, e.g., cdc25. • Cyclins form the regulatory subunits and CDKs the catalytic subunits of an activated heterodimer; cyclins have no catalytic activity and CDKs are inactive in the absence of a partner cyclin. When activated by a bound cyclin, CDKs perform a common biochemical reaction called phosphorylation that activates or inactivates target proteins to orchestrate coordinated entry into the next phase of the cell cycle. Different cyclin-CDK combinations determine the downstream proteins targeted. CDKs are constitutively expressed in cells whereas cyclins are synthesised at specific stages of the cell cycle, in response to various molecular signals.
Mitosis - meiosis • a | In mitosis, diploid cells replicate chromosomes during S phase and segregate sister chromatids during M phase, so that diploid daughter cells are produced. • b | In meiosis, two chromosome-segregation phases, meiosis I and meiosis II, follow a single round of DNA replication during pre-meiotic S phase. During meiosis I, homologous chromosomes (shown in red and blue) are segregated to opposite poles. Sister chromatids segregate to opposite poles during meiosis II, which results in the formation of non-identical haploid gametes. Please note that the lengths of the cell-cycle stages are not drawn to scale.
Cell death • Two cell death pathways, necrosis and apoptosis. Necrosis involves break-down of the cellular membrane, which leads to leakage of intracellular proteins to the extracellular space and subsequently, inflammation. Necrosis usually affects large groups of cells while apoptosis typically involves single cells that undergo organised destruction of the cellular cytoskeleton and formation of apoptotic bodies, which are phagocytosed without an inflammatory reaction. http://herkules.oulu.fi/isbn9514266676/html/i267388.html
Necrosis • Many diverse initiating conditions that trigger necrosis may provoke a net increase in the cytoplasmic calcium concentration, either by stimulating uptake of extracellular calcium or by facilitating the release of calcium stores from the endoplasmic reticulum. Calcium could, in turn, signal the mobilization of executioner cathepsin proteases and other hydrolases through calpain activation. Calpains have also been implicated in the activation of pro-apoptotic caspase proteases.
Apoptosis is a cellular response to a cellular "insult" such as UV light, chemical or physical damage or a viral infection. This insult starts a cascade of events which lead to the destruction of the cell. This mechanism is often called "programmed cell death" as it is an innate response of the cell which protects the rest of the organism from a potentially harmful agent.
Apoptosis • In an organism of a healthy person the cellular homeostasis is defined by a balance between the destructive and proliferative processes of cells. Apoptosis is a life long programmed, energetically dependent and genetically regulated cell destruction process. This cellular destruction process works through a special signal and gets rid of weak, unnecessary and damaged cells from an organism. Every day, approximately 5 % cells of an organism under go apoptosis, and their place occupy new cells. During this process the cells die away within 15-120 minutes without leaving any marks behind. TNF-a and Fas-ligand (CD178) switch on a cascade of biochemical reactions, whose final stage includes a chromosome defragmentation and consequently a complete loss of the cell. On the surface of cells present special receptors for TNF-a: TNF-RI (molecular weight- 55-60 kDa) and TNF-RII (molecular weight- 75-80 kDa), where as for a Fas-ligand exists a receptor so called Fas/APO-1 (CD95).
Apoptosis: cell morphology • The earliest indications of apoptotic cell death are morphological alterations of the cells such as chromatin condensation, disappearance of the nucleolus, and alterations of the cell surface, characterized by the occurrence of blebs. These signs are followed by a margination of the chromatin at the inner surface of the nuclear membrane. Eventually the activation of a variety of nucleases leads to the fragmentation of DNA. DNA degradation during apoptosis generally occurs at two levels: early as high molecular weight fragments and later on as fragments of the size of nucleosomes. This requires a number of specific DNases, which are activated specifically during apoptosis. DNA fragmentation can be used to identify apoptotic cells
Mitochondria in Apoptosis • Increases in cytosolic Ca2+ levels due to activation of ion channel-linked receptors, such as that for the excitatory amino acid neurotransmitter glutamic acid, can induce permeability transition (PT) of the mitochondrial membrane. PT constitutes the first rate-limiting event of the common pathway of apoptosis. Upon PT, apoptogenic factors leak into the cytoplasm from the mitochondrial intermembrane space. Two such factors, cytochrome c and apoptosis inducing factor (AIF), begin a cascade of proteolytic activity that ultimately leads to nuclear damage (DNA fragmentation, DNA mutations) and cell death. Cytochrome c, a key protein in electron transport, appears to act by forming a multimeric complex with Apaf-1, a protease, which in turn activates procaspase 9, and begins a cascade of activation of downstream caspases. Smac/Diablo is released from the mitochondria and inhibits IAP (inhibitor of apoptosis) from interacting with caspase 9 leading to apoptosis. Bcl-2 and Bcl-X can prevent pore formation and block the release of cytochrome c from the mitochondria and prevent activation of the caspase cascade and apoptosis. PT is also related to the mitochondrial generation of reactive oxygen species which plays a role in the degradation phase of apoptosis (i.e. plasma membrane alterations).
Lipid signaling in apoptosis • a Phosphatidylserine (PS) accumulates in the outer leaflet of the plasma membrane of apoptotic cells. • b Apoptotic neurons with condensed nuclei (stained blue with Hoechst 33342) and phosphatidylserine exposed on the cell surface, as visualized by staining with FITC (fluorescein isothiocyanate)–annexin V (green)140. • c Regulation of phosphatidyl-serine distribution between the two leaflets of the plasma membrane. Phospholipid scramblase (PLS) activity is Ca2+ dependent, whereas aminophospholipid trans-locase (APT) is inhibited by Ca2+ and SH-reagents (N-ethylmaleimide, NEM) and is vitally dependent on the intracellular ATP concentration. • d Normal viable cells are not engulfed by macrophages. Active repulsion of viable cells can be achieved by cell-surface molecules, which transmit detachment signals. Platelet–endothelial cell adhesion molecule-1 (PECAM-1, also known as CD31) normally prevents phagocyte recognition of viable cells. In apoptosis, the function of CD31 changes from repulsive (orange) to adhesive (green) and promotes phagocytosis. In some cells, CD31 engagement activates a cation-nonspecific channel, which mediates Ca2+ influx. Changes in the properties of CD31 in apoptotic cells might modulate Ca2+ signals and the subsequent exposure of phosphatidylserine and/or annexin I.