Download
1 / 27
320 likes | 1.01k Views
Lecture 20 Protein Targeting. The Rough ER: translocation and secretion reading: Chapter 13. Targeting of proteins: Location, location, location Constant growth required to maintain unique structure ...part of dissipative structure

E N D
Lecture 20 Protein Targeting The Rough ER: translocation and secretion reading: Chapter 13
Targeting of proteins: • Location, location, location • Constant growth required to maintain unique structure • ...part of dissipative structure • Organelles are not made de-novo…expand existing structures
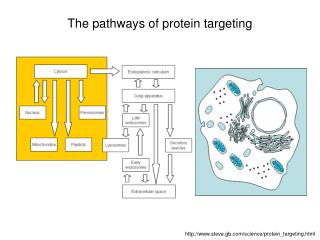
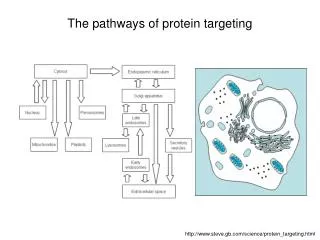
Secreted proteins are sequestered from the cytoplasm in microsomes
A 16-30 residue hydrophobic signal sequence directs the ribosome to the ER The hydrophobic core of the signal sequence contains one or more “+” charged residue
Rough ER • Targeting sequence on protein…not mRNA • Must be translated to be read • Translation must stop…wait to dock with site on RER (why?) • Signal-recognition particle, SRP, stops translation and provides for delivery …mailman…recognizes 3 structures
Bacterial Homologs: With GTP molecules bound FtsY and Ffh recognize each other. Cleavage of GRPs leads to complex disassembly
Sec61 (SecY) from Methannococcus jannaschii A ring of Isoleucine residues forms a hydrophobic ‘seal’ in the middle of the channel The ‘plug’ helix moves out during translocation Lateral exit into the bilayer can be permitted if blue helices separate
Liposome reconstitution experiments have demonstrated that Sec61, SPR receptor, and the nascent protein complexed with ribosome and SPR are absolutely required for translocation. No additional energy is required for translocation Can proteins be translocated not co-translationally, but post-translatonally?
Yes, in yeast successive binding of BiP-ADP makes transport unidirectional BiP is a chaperone protein The signal sequence is cleaved soon after (in both mechanisms)
Translocon is dynamic • Soluble proteins end up in “exoplasmic space” …topologically equivalent to the extracellular space • Transmembrane segments of membrane proteins move sideways into RER membrane
Translocation of type I membrane proteins require (1) signal sequence (cleaved) and (2) stop-transfer anchor sequence
Type II Type III Both of these protein types require just one internal hydrophobic signal-anchor sequence
Threading membrane proteins through the membrane Type I…targeting sequence and stop-transfer sequence ….SRP/receptor…cleaved sequence Type II, III…internal sequences…oriented by positive cluster…why? Type IV…multipass…target…stop transfer…etc.
Why anchors are positive (+++)? Membrane interior always has positive potential inside, it would repel +++ clusters most effectively membrane +400 to +700 mV Electric potential dipoles created by lipid carbonyls and oriented water
GPI anchored proteins Purpose? …Lateral diffusion….? …Targeting? Transfer of type 1 protein
Hydrophobicity profiles of primary sequences GLUT1 protein
N-linked poly-sugar chains are synthesized on Dolichol phosphate utilized as an attachment anchor Dolicol = 75-90 carbon isoprenoid lipid
Oligosacharides attached to proteins help folding through specific associations with lectins Removal and re-attachment of one glucose residue acts as a ‘quality control’ step in the process of folding (see text).
Proper folding of Hemagglutiin (HAo) occurs in the presence of chaperones (Bip) and two types of lectins (Calnexin and Calreticulin). The folded structure works as a pre-loaded spring in the mechanism of Influenza virus membrane fusion mediated by HAo
Oxidized disulfide Reduced dithiol form PDI = protein disulfide isomerase
What if the protein has more than one disulfide bond? DPI = protein disulfide isomerase
Proper folding of secreted protein • formation of correct S-S bonds • facilitate slow steps… peptidilyl-prolyl-isomerase (cyclophilin) • …cyclosporin A …tissue rejection