Download
1 / 1
10 likes | 98 Views
Molecular evolution of nrt2 , a nitrate transporter gene, with an emphasis on Hebeloma phylogeny. Jason C. Slot and David S. Hibbett Department of Biology, Clark University, Worcester, MA 01610 Jslot@clarku.edu , dhibbett@clarku.edu.

E N D
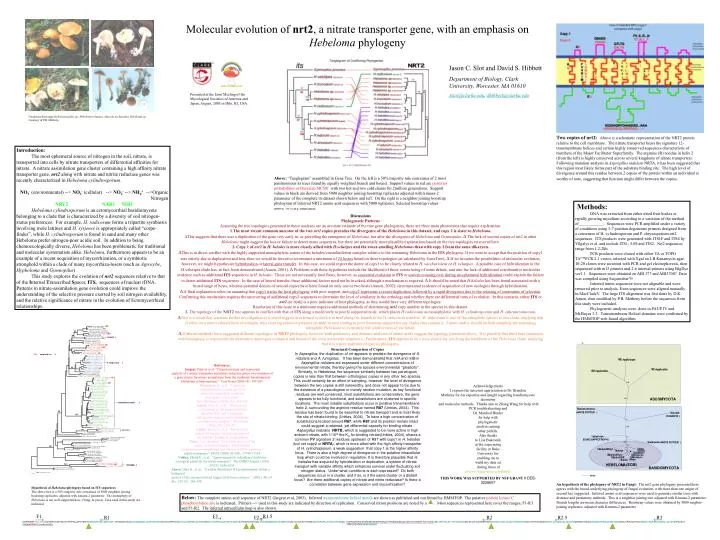
Molecular evolution of nrt2, a nitrate transporter gene, with an emphasis on Hebeloma phylogeny Jason C. Slot and David S. Hibbett Department of Biology, Clark University, Worcester, MA 01610 Jslot@clarku.edu, dhibbett@clarku.edu Presented at the Joint Meeting of the Mycological Societies of America and Japan, August, 2005 in Hilo, HI, USA Clockwise from upper left Gymnopilus sp., Hebeloma velutipes, Alnicola escharoides, Hebeloma sp. Courtesy of P.B. Matheny Two copies of nrt2:Above is a schematic representation of the NRT2 protein relative to the cell membrane. The nitrate transporter bears the signature 12-transmembrane helices and certain highly conserved sequences characteristic of members of the Major Facilitator Superfamily. The arginine (R) residue in helix 2 (from the left) is highly conserved across several kingdoms of nitrate transporters. Following mutation analysis in Aspergillus nidulans NRTA, it has been suggested that this region most likely forms part of the substrate binding site. The high level of divergence around this residue between 2 copies of the protein within an individual is worthy of note, suggesting that function might differ between the copies. Introduction: The most ephemeral source of nitrogen in the soil, nitrate, is transported into cells by nitrate transporters of differential affinities for nitrate. A nitrate assimilation gene cluster containing a high affinity nitrate transporter gene, nrt2 along with nitrate and nitrite reductase genes was recently characterized in Hebeloma cylindrosporum. NO3- (environmental) --> NO3- (cellular) --> NO2- --> NH4+ -->Organic Nitrogen NRT2 NAR1 NIR1 Hebeloma cylindrosporum is an ectomycorrhial basidiomycete belonging to a clade that is characterized by a diversity of soil nitrogen-status preferences. For example, H. radicosum forms a tripartite symbiosis involving mole latrines and H. syrjense is appropriately called “corpe-finder”, while H. cylindrosporum is found in sand and many other Hebeloma prefer nitrogen-poor acidic soil. In addition to being chemoecologically diverse, Hebeloma has been problematic for traditional and molecular systematists alike. Hebeloma, furthermore appears to be an example of a recent acquisition of mycorrhization, or a symbiotic stronghold within a clade of many mycorrhiza-losers (such as Agrocybe, Hypholoma and Gymnopilus). This study explores the evolution of nrt2 sequences relative to that of the Internal Transcribed Spacer, ITS, sequences of nuclear rDNA. Patterns in nitrate-assimilation gene evolution could improve the understanding of the selective pressure exerted by soil nitrogen availability, and the relative significance of nitrate in the evolution of Ectomycorrhizal relationships. Above: “Tanglegram” assembled in Gene Tree. On the left is a 50% majority rule concensus of 2 most parsimonious its trees found by equally weighted branch and bound. Support values in red are posterior probabilities of Bayesian MCMC with two hot and two cold chains for 2million generations. Support values in black are derived from 5000 neighbor joining bootstrap replicates adjusted with kimura-2 parameter of the complete its dataset shown below and left. On the right is a neighbor joining boostrap phylogram of inferred NRT2 amino acid sequences with 5000 replicates. Selected bootstrap values above 70% are indicated. Methods: DNA was extracted from either dried fruit bodies or rapidly growing mycelium according to a variation of the method of____________. Sequences were PCR amplified under a variety of conditions using 3-7 position degenerate primers designed from a consensus of H. cylindrosporum and P. chrysosporium nrt2 sequences. ITS products were generated with ITS1F and ITS4 by Vilgalys et al. and include ITS1, 5.8S and ITS2. Nrt2 sequences range from 1.2-2kb. PCR products were cloned with either TA or TOPO TA"℠PCR 2.1 vector, selected with Xgal on LB Kanamycin agar. 10-20 clones were screened with PCR and gel electrophoresis, then sequenced with m13 primers and 2-6 internal primers using BigDye ver1.1. Sequences were obtained on ABI 377 and ABI3700? Data was compiled using Sequencher"℮ Inferred intron sequences were not alignable and were removed prior to analysis. Exon sequences were aligned manually in MacCladeV. The large ITS alignment was first done by D.K. Aanen, then modified by P.B. Matheny before the sequences from this study were included. Phylogenetic analyses were done in PAUP IV and Mr.Bayes 3.3. Transmembrane Helical domains were confirmed by the HMMTOP web-based algorithm. Discussions Phylogenetic Patterns Assuming the tree topologies generated in these analyses are an accurate estimate of the true gene phylogenies, there are three main phenomena that require explanation. 1.The most recent common ancestor of the two nrt2 copies precedes the divergence of the Hebeloma in this dataset, and copy 2 is sister to Hebeloma. ∆This suggests that there was a duplication of the gene very early in, or preceding the emergence of Hebeloma, but after the divergence of Hebeloma and Gymnopilus. ∆ The lack of second copies of nrt2 in other Hebeloma might suggest the loss or failure to detect many sequences, but there are potentially more plausible explanations based on the tree topologies recovered here. 2.Copy 1 of nrt2 in H. helodes is more closely allied with H.velutipes and the sweet-smelling Hebelomas than with copy 2 from the same dikaryon. ∆This is in direct conflict with the highly supported monophyletic nature of the helodes/crustuliniforme complex relative to the remaining Hebeloma in the ITS phylogeny. If we were to accept that the position of copy1 was strictly due to duplication and loss, then we would be forced to reconstruct a minimum of 24 losses based on these topologies (as calculated by GeneTree). ∆ If we broaden the possibilities of molecular evolution, however, we might hypothesize hybridization or lateral gene transfer. In this case, we could expect the donor of copy1 to be closely related to H. sacchariolens or H. velutipes. A possible case of hybridization in the H.velutipes clade has, in fact, been demonstrated (Aanen, 2001). ∆ Problems with these hypotheses include the likelihood of these events being of some debate, and also the lack of additional corroborative molecular evidence such as additional ITS sequences in H. helodes. These are not necessarily fatal flaws, however, as concerted evolution in ITS or uneven crossing over during an ephemeral hybridization could explain the failure to detect additional ITS sequences. In the case of lateral transfer, these additional factors need not be invoked, although a mechanism is required. ∆ It should be noted that H.helodes has been found associated with a broad range of hosts, whereas potential donors of second copies have been found on only one or two hosts (Aanen, 2002); circumstantial evidence of acquisition of new ecologies through hybridization. ∆ A final explanation relies on assuming that copy1 tracks the host phylogeny with poor support, and copy2 represents a recent duplication followed by a rapid divergence due to the relaxing of constraints of selection. Confirming this mechanism requires the uncovering of additional copy2 sequences to determine the level of similarity in the orthologs and whether there are differential rates of evolution. In this scenario, either ITS or nrt2 (or both) is a poor indicator of host phylogeny, as they would have very different topologies. Resolution of this issue at a minimum requires additional methods of determining nrt2 copy number in the species in this dataset. 3. The topology of the NRT2 tree appears to conflict with that of ITS along a moderately to poorly supported node, which places H.radicosum as monophyletic with H. cylindrosporum and H. edurum/sinuosum. ∆This is a result that warrants further investigation as it could suggest accelerated evolution in nrt2 along the branch to the H.radicosum terminal. H. radicosum is one of the nitrophilic species in this clade, implying that it relies on a more reduced form of nitrogen, thus relaxing selective pressure on nrt2, in turn leading to poor bootstrap support for any clades that contain it. Future studies should include sampling the remaining nitrophilic Hebeloma to eventually test relative rates of evolution. ∆ Different methods have suggested different topologies in NRT2 phylogeny, however both parsimony and distance analyses of amino acids suggest the topology presented above. It is possible that third base saturation with homoplasy is responsible for alternative topologies in branch and bound of the exon nucleotide sequences. Furthermore, ITS appears to be a poor choice for resolving the backbone of the Hebeloma clade, implying that it is a poor indicator of species phylogeny. Structural Comparison of Copies In Aspergillus, the duplication of nrt appears to predate the divergence of A. nidulans and A. fumigatus. It has been demonstrated that nrtA and nrtB in Aspergillus nidulans are expressed under different concentrations of environmental nitrate, thereby giving the species environmental “plasticity”. Similarly, in Hebeloma, the sequence similarity between two paralogous copies is less than that between orthologous copies in any other two species. This could certainly be an effect of sampling, however the level of divergence between the two copies is still noteworthy, and does not appear to be due to the existence of a pseudogene or merely random mutation, as key functional residues are well conserved, most substitutions are conservative, the gene appears to be fully functional, and substitutions are clustered in specific locations. The most notable substitutions occur in putative transmembrane helix 2, surrounding the arginine residue named R87 (Unkles, 2004). This residue has been found to be essential to nitrate transport and is most likely the site of nitrate binding (Unkles, 2004). To have a high concentration of substitutions located around R87, while R87 and its position remain intact could suggest a retained, yet differential capacity for binding nitrate. Aspergillus nidulansNRTB, which is suggested to be more active in high ambient nitrate, with 1/10th the Km for binding nitrate(Unkles, 2004), shares a common FV signature 2 residues upstream of R87 with copy1 in H. helodes (but not copy2 or NRTA), which is more allied with the high affinity transporter of H. cylindrosporum, a weak suggestion that copy 1 is the higher affinity locus. There is also a high degree of divergence in the putative intracellular loop which could be involved in regulation. It is therefore plausible that H. helodes has acquired by hybridization or duplication, a system of nitrate transport with variable affinity which enhances survival under fluctuating soil nitrogen status. Under what conditions is each expressed? Do both sequences occur in a cluster, and if so, is it the same cluster or a distant locus? Are there additional copies of nitrate and nitrite reductase? Is there a correlation between gene expression and mycorrhization? References: Jargeat, Patricia et al. “Characterixation and expression analysis of a nitrate transporter and nitrate reductase genes, two members of a gene cluster for nitrate assimilation from the symbiotic basidiomycete Hebelomacylindrosporum.” Curr Genet (2003) 43: 199-205. Marmeisse, R., et al. “Hebeloma cylindrosporum-a model species to study ectomycorrhizal symbiosis from gene to ecosystem.” New Phytologist (2004) 163: 481-498. Aanen, Duur K., et al. “Phylogenetic relationships in the genus Hebeloma based on ITS1 and 2 sequences, with special emphasis on the Hebeloma crustuliniforme complex.” Mycologia (2002) 92(2):269-281. Unkles, Sheila E., et al. “Two perfectly conserved arginine residues are required for substrate binding in a high-affinity Yang, et al., (in press) “New Asian species of the genus Anamika (euagarics, hebelomatoid clade) based on morphology and ribosomal DNA sequences.” nitrate transporter.” PNAS (2004) 101(50): 17549-17554. Unkles, Shelia E., et al., “Apparent genetic redundancy facilitates ecological plasticity for nitrate transport.” The EMBO Journal (2001) 20(22): 6246-6255. Aanen, Duur K., et al., “A widely distributed ITS polymorphism within a biological species of the ectomycorrhizal fungus Hebeloma velutipes.” (2001). Mycol. Res. 105 (3): 284-290 Acknowledgements I express the sincerest appreciation to Dr. Brandon Matheny for his expertise and insight regarding basidiomycete taxonomy and molecular methods. Thanks also to Zhang Wang for help with PCR troubleshooting and Dr. Manfred Binder for help with phylogenetic analysis among other pitfalls. Also thanks to Lisa Bukovnik at the sequencing facility at Duke University for enabling me to build my data set during times of adverse sequencing conditions. THIS WORK WAS SUPPORTED BY NSF GRANT # DEB-0228657 An hypothesis of the phylogeny of NRT2 in Fungi: The nrt2 gene phylogeny presented here agrees with the broad, underlying phylogeny of fungal evolution, with more than one origin of second loci suggested. Inferred amino acid sequences were used to generate similar trees with distance and parsimony methods. This is a neighbor joining tree adjusted with Kimura-2 parameter. Branch lengths are mean character differences. Bootstrap values were obtained by 5000 neigbor-joining replicates, adjusted with Kimura-2 parameter. Hypothesis of Hebeloma phylogeny based on ITS sequences The above tree is a 50% majority rule consensus of 5000 neighbor joining bootstrap replicates, adjusted with kimura-2 parameter. The monophyly of Hebeloma is not well supported here. (Yang, in press) Taxa used in this study are indicated. Below:The complete amino acid sequence of NRT2 (Jargeat et al, 2003). Inferred transmembrane helical motifs are shown as published and confirmed by HMMTOP. The putative protein kinase C phosphorylation site is indicated. Primers --> used in this study are indicated by direction of replication. Conserved intron positions are noted by a . Most sequences represented here cover the ranges, F1-R3 and F1-R2. The inferred intracellular loop is also shown. F1 F3 R1.5 R1 F2 R2 R2.5 R3 MSPPKSSRGAPKFKWSHLWEPAIVNPVNLKSYTIPIFNLGDPYARAFHLSWLGFFVAFLSWFAFPPLIPDAIKSDLHLSAAQVANSNIIALCATFVVRVGVGPLVDQYGPRKVMAYLLILGAIPSGLAGTARSAEGLYVLRFFIGILGATFVPCQAWTSAFFDKNCVGTANALVGGWGNMGGGATFAIMTSLFQSLTQTYGLSTHVAWRAAFAIVPAPILLFVAVLTFIFGQDHPAGKWSERHTLPVAALAVQQG HQIPNIQPEKSLKSSSESSKDEKDPEGNAAVTVRPAIADEDLALVKSTVDVAINEPLTLKTTVKILTNPLTWLPALAYLTTFGVELAIDSKFADVLFVLFSKRRPGFDQTTAGYYTSILGLLNLVTRPAGGYFGDLVYRHYGTNGKKAWTLLCGLIMGAALVAGGFYMQNNRTSGDEQLSVLMGVFSVAAIFSEFGNGANFALVPHCNAYNNGVMSGLVGSFGNLGGIIFALVFRFQTEVGKAFWIMGVISIGIN ALLIPVSVPAL(521)




















