Download

1 / 20
210 likes | 367 Views
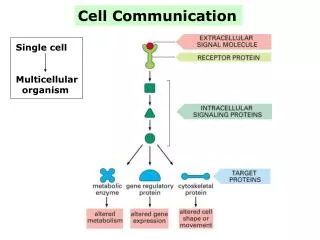
Vav Deficient Macrophages: The Roles of Rac and Rho in Cell Migration. Vav-Vav-voom, let’s Rac ’n’ Rho! Julia Sero Ty Thomson 12/10/2002. Models of Macrophage Migration. Hematopoietic cells migrate during development and immune response Macrophage migration mechanism is not well understood

E N D
Vav Deficient Macrophages: The Roles of Rac and Rho in Cell Migration Vav-Vav-voom, let’s Rac ’n’ Rho! Julia Sero Ty Thomson 12/10/2002
Models of Macrophage Migration • Hematopoietic cells migrate during development and immune response • Macrophage migration mechanism is not well understood • Genetic knockout is a molecular biological tool to investigate gene product functions • Modeling is an engineering tool to help direct research and verify hypotheses
Vav- Cell Phenotype • Vav: guanine nucleotide exchange factor and adaptor protein (3 isoforms) • Vav1>Vav3>>Vav2 • Activates Rho GTPases (Rac, Rho, Cdc42) • Involved in signal transduction from integrins and other receptors via multiple pathways • Vav-deficient macrophages show migration defect • Vav1-/-;Vav3-/- extend multiple lamellipodia but do not migrate in wound healing assay • Short, narrow lamellipodia • Fail to translocate nuclei or retract tails
Movies: Wild type: Double knock out:
Vav pathways Integrin Src Syk Active Vav F-actin Rac Rho F-actin MAPK/ERK pathway ROK Calpain Activation Myosin Phosphorylation
Hypothesis • Local signals mediated by Vav complexes crucial for downstream effects • Rac versus Rho type adhesions: • Dynamic and propulsive (calpain) • Stable and anchoring (myosin contraction) • Balance of signal outputs is necessary for efficient migration • DKO cells have an imbalance between focal adhesion turnover and intracellular contraction due to loss of Vav
MAPK Pathway (Modified from Bhalla et al., 2002)
Myosin Activation Pathway Active Vav GDP-Rho GTP-Rho GTP-Rho/ ROK ROK MLC MLC* GTP-Rho/ MBS MBS
Major Model Assumptions • Vav1, Vav2 and Vav3 all have the same activity • Localized signaling can studied, and the effects summed to account for overall behavior
Differential of Contraction and Focal Adhesion Turnover Physiological Range? Pathological Range?
Summary of Observations • Calpain production and activation only significant for active Vav concentrations of greater than ~1nM • Myosin activation significant for Vav concentrations of greater than ~0.1nM • Postulated wild type active Vav concentration at about 10-100nM • If [Vav2]/[Vav1+Vav3] = (1/10 to 1/100), then DKO could fall in region of myosin activation but low calpain activation
Model Predictions • Normal active Vav concentrations in the range of 10-100nM, while active Vav2 in DKO concentration about 0.4-4nM • Vav DKO macrophage phenotype may be as result of imbalance between focal adhesion turnover and myosin mediated contraction
Other Considerations • Vav also interacts with Cdc42, which is thought to be important for cell polarity • Rac and Rho are also involved in F-actin polymerization • Time delay observed between Rac and Rho activation and maturation of focal contacts • Signaling upstream of Vav has been ignored • Other pathways downstream of Vav affected • Concentrations of Rac, Rho, ROK, MLC, MBS all assumed to be 0.2uM
Proposed Experiments • Observe DKO phenotype on other substrates in response to different signals • Measure biochemical interactions between species (kinetics) in vitro • Assay for myosin phosphorylation and active calpain • Tension force assay to determine if DKOs lack propulsive traction forces at leading edge • Measure half life of focal adhesions (GFP-tagged proteins, confocal microscopy)
Thank you! • Joan Brugge, Amy Hall, and the Brugge lab (Dept. of Cell Biology, Harvard Medical School) • Reshma Shetty • Ali Khademhosseini • Doug Lauffenburger
References • Abe K., et al. Vav2 is an activator of Cdc42, Rac1, and RhoA. J. Biol. Chem. 257(14):10141-10149. 2000. • Amano, M., et al. Phosphorylation and activation of myosin by Rho-associated kinase. J. Biol. Chem. 271(34):20246-20249. 1996. • Astagarthi A.R., Nelson C.M., Horwitz A.F., Lauffenburger D.A. Quantitative relationship among integrin-ligand binding, adhesion, and signaling via focal adhesion kinase and extracellular signal-regulated kinase 2. J. Biol. Chem. 274(38):27119-27127. 1999. • Ballestrem C., Hinz B., Imhof B.A., Wehrle-Haller B. Marching at the front and dragging behind: differential V3-integrin turnover regulates focal adhesion behavior. J. Cell Biol. 155(7):1319-1332. 2001. • Beningo K.A., Dembo M., Kaverina I., Small V.J., Wang Y. Nascent focal adhesions for the generation of strong propulsive forces in migrating fibroblasts. J. Cell Biol. 153(4):881-887. 2001. • Bhalla U.S. , Ram P.T., Iyengar R. MAP kinase phosphatase as a locus of flexibility in a mitogen-activated protein kinase signaling network. Science. 297:1018-1023. 2002. • Bustelo, X.R. Regulatory and signaling properties of the Vav family. Mol. Cell Biol. 20(5):1461-1477. 2000. • Carragher N.O., Frame M.C. Calpian: a role in cell transformation and migration. Int. J. Biochem. Cell Biol. 1322:1-5. 2002. • Cox E.A. and Huttenlocher A. Regulation of integrin-mediated adhesion during cell migration. Microsc. Res. Tech. 43:412-419. 1998. • DiMilla P.A., Barbee K., Lauffenburger D.A. Mathematical model for the effects of adhesion and mechanics on cell migration speed. Biophys. J. 60:15-37. 1991. • Eblen S.T., Slack J.K., Weber M.J., Catling A.D.Rac-PAK signaling stimulates extracellular signal-regulated kinase (ERK) activation by regulating formation of MEK1-ERK complexes. Mol. Cell Biol. 22(17):6023-6033. 2002 • Giniger E. How do Rho family GTPases direct axon growth and guidance? A proposal relating signaling pathways to growth cone mechanics. Differentiation. 70:385-396. 2002. • Glading A., Uberall F., Keyse S.M., Lauffenburger D.A. Membrane proximal ERK signaling is required for M-calpain activation downstream of epidermal growth factor receptor signaling. J. Biol. Chem. 276(26):23341-23348. 2001. • Glading, A., Lauffenburger D., Wells A. Cutting to the chase: calpain proteases in cell motility. TRENDS in Cell Biol. 12(1):46-54. 2002. • Jones G.E., Allen W.E., Ridley A.J. The Rho GTPases in macrophage motility and chemotaxis. Cell Adhes. Commun. 6(2-3):237-245. 1998. • Kulkarni S., Goll D., Fox, J.E.B. Calpain cleaves RhoA generating a dominant-negative form that inhibits integrin-induced actin filament assembly and contraction. J. Biol. Chem. 277(27):24435-24441. 2002. • Lauffenburger D.A. and Horwitz A.F. Cell migration: a physically integrated molecular process. Cell. 84(3):359-69. 1996. • Miranti C.K., Leng L., Maschberger P., Brugge J.S., Shattil S.J. Identification of a novel integrin signaling pathway involving the kinase Syk and the guanine nucleotide exchange factor Vav1. Curr. Biol. 8(24):1289-99. 1998. • Moores S.L., Selfors L.M., Fredericks J., Breit T., Fujikawa K., Alt F.W., Brugge J.S., Swat W. Vav family proteins couple to diverse cell surface receptors.Mol. Cell Biol. 20(17):6364-73. 2000. • Nobes C.D. and Hall A. Rho GTPases control polarity, protrusion, and adhesion during cell movement. J. Cell Biol. 144(6):1235-1244. 1999. • Orkin S. and Zon L. Hematopoiesis and stem cells: plasticity versus developmental heterogeneity. Nat. Immunol. 3(4):323-8. 2002. • Petit V. and Thiery J. Focal adhesions: structure and dynamics. Biol. Cell. 92:477-494. 2000. • Schoeber B., Eichler-Jonsson C., Gilles E.D., Muller G. Computational modeling of the dynamics of the MAP kinase cascade activated by surface and internalized EGF receptors. Nature Biotechnol. 20:370-375. 2002. • Turner M. and Billadeau D.D. Vav proteins as signal integrators for multi-subunit immune-recognition receptors. Nature Reviews Immunol. 2:476-486. 2002. • Wang H.B., Dembo M., Hanks S.K., Wang Y. Focal adhesion kinase is involved in mechanosensing during fibroblast migration. P.N.A.S. USA. 98(20):11295-300. 2001. • Wehrle-Haller B. and Imhof B.A. The inner lives of focal adhesions. TRENDS in Cell Biol. 12(8):382-389. 2002. • Worthylake R.A., Lemoine S., Watson J.M., Burridge K. RhoA is required for monocyte tail retraction during transendothelial migration. J. Cell Biol. 2001. 154(1):147-160 • Yeung Y., Wang Y., Einstein D.B., Lee P.S.W., Stanley E.R. Colony-stimulating factor-1 stimulates the formation of multimeric cytosolic complexes of signaling proteins and cytoskeletal components in macrophages. J. Biol. Chem. 273(27):17128-17137. 1998. • Yoshinaga-Ohara N., Takahashi A., Uchiyama T., Sasada M. Spatiotemporal regulation of moesin phosphorylation and rear release by Rho and serine/threonine phosphatase during neutrophil migration. Exp. Cell Res. 278(1):112-122. 2002. • Zeng L., Sachdev P., Yan L., Chan J.L., Trenkle T., McClelland M., Welsh J., Wang L.H. Vav3 mediates receptor protein tyrosine kinase signaling, regulates GTPase activity, modulates cell morphology, and induces cell transformation. Mol. Cell Biol. 20(24):9212-24. 2000.