Download
1 / 19
190 likes | 404 Views
Eukarüootne transkriptsioon. RNA polümeraasid. Kõikidel eukarüootidel on kolm erinevat RNA polümeraasi, mis täidavad erinevaid ülesandeid. Kõik RNA polümeraasid on suured kompleksid koosnedes paljudest erinevatest valkudest.

E N D
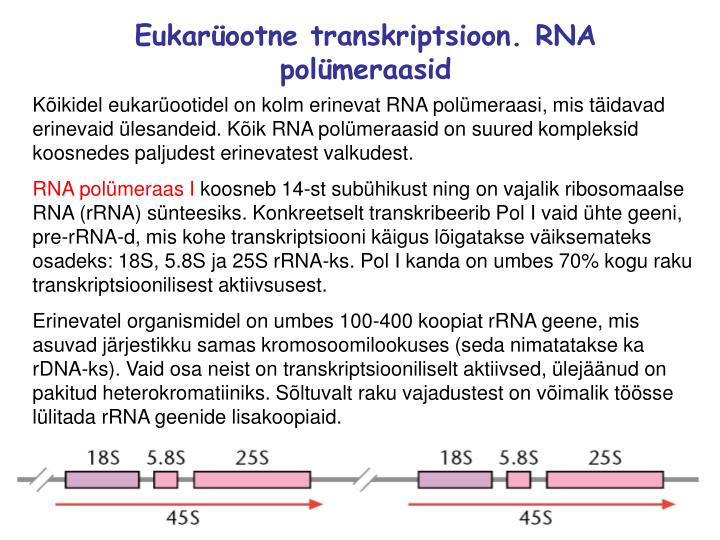
Eukarüootne transkriptsioon. RNA polümeraasid Kõikidel eukarüootidel on kolm erinevat RNA polümeraasi, mis täidavad erinevaid ülesandeid. Kõik RNA polümeraasid on suured kompleksid koosnedes paljudest erinevatest valkudest. RNA polümeraas I koosneb 14-st subühikust ning on vajalik ribosomaalse RNA (rRNA) sünteesiks. Konkreetselt transkribeerib Pol I vaid ühte geeni, pre-rRNA-d, mis kohe transkriptsiooni käigus lõigatakse väiksemateks osadeks: 18S, 5.8S ja 25S rRNA-ks. Pol I kanda on umbes 70% kogu raku transkriptsioonilisest aktiivsusest. Erinevatel organismidel on umbes 100-400 koopiat rRNA geene, mis asuvad järjestikku samas kromosoomilookuses (seda nimatatakse ka rDNA-ks). Vaid osa neist on transkriptsiooniliselt aktiivsed, ülejäänud on pakitud heterokromatiiniks. Sõltuvalt raku vajadustest on võimalik töösse lülitada rRNA geenide lisakoopiaid.
Eukarüootne transkriptsioon. RNA polümeraasid RNA polümeraas II koosneb 12-st subühikust ning on vastutav kogu mRNA sünteesi eest. Lisaks sellele transkribeerib Pol II ka mitmeid väikesi tuuma RNA-sid (snRNA – small nuclear RNA). Pol II geene on küll kõige rohkem, kuid kogu rakus toimuvast transkriptsioonist vaid umbes 20% on teostatud Pol II poolt. RNA polümeraas III koosneb 17-st subühikust. Pol III sünteesib kõik tRNA-d ja lisaks ka 5S rRNA ning enamuse väiksemaid mittetransleeritavaid RNA-sid (7SK, MRP, U6, H1 ja teised, valdavalt regulatoorse tähtsusega RNA-d). Kokku langeb Pol III osaks umbes 10% rakus toimuvast transkriptsioonist. Tänu rRNA ja tRNA suuremale stabiilsusele, moodustavad need raku totaalsest RNA-st umbes 95%. Kõige paremini on uuritud Pol II sõltuvat transkriptsiooni.
Basaalsed transkriptsioonifaktorid. Kasutatakse nii mõisteid basal transcription factors kui ka general transcription factors (GTF). Need on valgud või valgukompleksid, mis on lisaks RNA polümeraasile vajalikud transkriptsiooni toimumiseks. Erinevad polümeraasid võivad vajada erinevaid faktoreid, seetõttu tähistatakse neid vastavalt polümeraasile, mille juurde nad kuuluvad. Näiteks TFIID (transcription factor II D) on vajalik Pol II sõltuvaks transkriptsiooniks ning TFIIIC on vajalik Pol III transkriptsiooniks. Transkriptsiooni toimumiseks peab RNA polümeraas seonduma geeni promootorile, selle avama ning alustama RNA sünteesi, seda nimetatakse transkriptsiooni initsiatsiooniks. Initsiatsiooni toimumiseks peab esmalt moodustuma promootoril pre-initsiatsiooni kompleks (PIC – pre-initiation complex). Pole lõpuni selge, kas PIC seondub promootorile korraga ühe suure kompleksina, või toimub selle moodustumine järk-järguliselt. Üldiselt eelistatakse mudelit, kus PIC moodustub promootoril mitme teineteisele järgneva etapi jooksul.
Preinitsiatsiooni kompleksi moodustumine Asi algab TFIID seondumisega promootorile. TFIID koosneb valgust TBP (TATA binding protein) ja sellega seondunud valkudest TAF-idest (TBP associated factors). Erinevaid TAF-e on vähemalt 13, kuid üldiselt võib TFIID koostis olla erinevates olukordades erinev, seda just TAF-ide arvel. TBP eelistab seonduda DNA järjestusele TATAA ning tema seondumisega kaasneb DNA ahela käändumine. Samas seondub TBP ka nendele promootoritele, mis TATA-box-i ei sisalda.
Preinitsiatsiooni kompleksi moodustumine Mõnede TAF-ide struktuur on väga sarnane histoonide struktuuriga. See annab alust oletada, et promootorile seondunult võib TFIID siduda DNA-d sarnaselt nukleosoomiga.
Preinitsiatsiooni kompleksi moodustumine TFIID on üks peamisi komponente millega seonduvad transkriptsiooni aktiveerivad valgud. TAF1 (TAFII250) omab histoonide atsetüültransferaasi aktiivsust ning samas kuulub osa TAF-e ka mõnede teiste HAT komplekside (näiteks SAGA) koosseisu, mistõttu piir TFIID ja kromatiini modifitseerivate komplekside vahel on üsna tinglik - edukaks transkriptsiooni aktivatsiooniks tuleb kohale tuua nii HAT kompleks kui ka TFIID. Peale TFIID-d seonduvad PIC-iga TFIIA ja TFIIB. TFIIA koosneb kolmest subühikust ning on vajalik eesätt TFIID seondumise stabiliseerimiseks ning TFIID-ga konkureerivate repressorvalkude tõrjumiseks. TFIIB on üks valk ning see seondub nii TBP kui DNA-ga ja on vajalik nii TFIID seondumise stabiliseerimiseks kui ka promootori “suuna” määramisel. Sageli on TATA-box-e ees veel teinegi spetsiifiline DNA järjestus BRE (TFIIBrecognition element). TFIIB on vajalik RNA polümeraasi seondumiseks PIC-iga.
Preinitsiatsiooni kompleksi moodustumine Järgmisena seondub PIC-iga RNA polümeraasi kompleks koos TFIIF-ga. Samuti on RNA polümeraasiga seondunud Mediator kompleks. TFIIF koosneb kahest subühikust ning on koos TFIIB-ga vajalik RNA polümeraasi seondumiseks PIC-ga. Mediaator on suur valgukompleks, koosneb 20-30 subühikust. Mediaator on samuti väga sageli transkriptsiooni aktivaatorite sihtmärgiks: püüdes kinni Mediaatori, saab promootorile tuua ka RNA polümeraasi. Mediaator ongi vahelüli transkriptsiooni aktivaatorite ja RNA polümeraasi vahel.
Preinitsiatsiooni kompleksi moodustumine Järgmisena seonduvad PIC-ga TFIIE ja TFIIH. TFIIE seondub tetrameerina, koosnedes 2 x 2 subühikust. TFIIE on vajalik RNA polümeraasi PIC-ile seondumise stabiliseerimiseks (tõenäoliselt aidates polümeraasil DNA-ga seonduda) ning samuti on TFIIE vajalik transkriptsiooni üleminekul elongatsioonifaasi. TFIIE aitab ka TFIIH-l seonduda PIC-iga ning TFIIE puudumisel ei suuda TFIIH promootorit avada. TFIIH koosneb kümnest subühikust ning seda võib jaotada tinglikult kaheks erinevaks kompleksiks: core complex (6 subühikut) on DNA helikaas, CAK kompleks (3 subühikut) on kinaas. TFIIH on roll transkriptsiooni initsiatsioonil on avada DNA ahelad (TFIIH on helikaas), mis lubab RNA polümeraasil alustada RNA sünteesi. CAK sub-kompleksi roll on fosforüleerida RNA polümeraasi suurima subühiku C-terminaalset domääni (CTD), mis on vajalik polümeraasi vabanemiseks PIC-ilt ning transkriptsiooni üleminekul elongatsioonifaasi. TFIIH on seotud märksa rohkemaga kui transkriptsiooni initsiatsioon. TFIIH on ka transkriptsiooni elongatsioonifaktoriks ning osaleb ka DNA reparatsioonil.
Preinitsiatsiooni kompleksi moodustumine Pol I ja Pol III promootoritel Pol I ja Pol III promootoritel moodustub PIC üldjoontes sarnaselt Pol II promootoritega, kuid erinevaid faktoreid on vähem. Pol I: koos seonduvad UBF (upstream binding factor) ja SL1 (promoter selectivity factor). SL1 koosneb TBP-st ja kolmest TAF-ist. TIFIA (transcription intermediary factor) seondub SL1 ja Pol I-ga. Pol I seondub ka veel vähemalt kahe faktoriga TIFIC ja TFIIH. Pol III promootorid on reeglina transkribeeritava ala sees. Promootorit tunneb ära TFIIIC (5 subühikut), mis toob kohale TFIIIB (koosneb TBP-st ja kahest TAF-ist) TFIIIB toob kohale Pol III.
Promootori vabanemine Kui PIC on koos, siis TFIIH (mis on DNA helikaas) alustab DNA ahelate avamist promootoril (moodustub nn “avatud kompleks” – open complex) ning peale seda alustab polümeraas RNA sünteesi. Transkriptsiooni täpne alguskoht pole kindlalt fikseeritud, see asub tavaliselt umbes 30-45 aluspaari pärast TATA-boxi, kuid mõnel juhul võib see olla isegi 100 aluspaari kaugusel. Tavaliselt ei lähe transkriptsioon käima kohe esimesel katsel. Reeglina sünteesitakse korduvalt umbes 3-10 nukleotiidist koosnevaid RNA lõike, enne kui alustatakse täispika mRNA sünteesi. Kui RNA pole vähemalt 10 nukleotiidi pikk, siis DNA-RNA-polümeraasi kolmikkompleks pole veel piisavalt stabiilne ning polümeraas võib väga kergesti RNA sünteesi katkestada. See on olukord kus transkriptsiooni initsiatsioon läheb üle elongatsiooniks ning sellega kaasnevad muudatused PIC-i struktuuris. Toimub promootori vabanemine (promoter clearance), et saaks toimuda järgmise PIC-i moodustumine. Sisuliselt tähendab see PIC-i komponentide jagunemist kaheks osaks: need mis jäävad promootorile ja need mis liiguvad RNA polümeraasiga kaasa. Samuti tomub polümeraasi seondumine transkriptsiooni elongatsiooniks ning mRNA protsessinguks vajalike valkudega. Efektiivseks elongatsiooniks peab polümeraas vabanema näiteks TFIIB-st.
RNA polümeraas II Koosneb 12-st subühikust (Rpb1-12), millest suurimad on Rpb1 ja Rpb2. Rpb1 CTD (C-terminal domain) on koht, kuhu seondub väga arvukalt erinevaid transkriptsiooniga kaasnevaid faktoreid. CTD interaktsioone reguleeritakse selle fosforüleerimiste kaudu.
Table 1 S. cerevisiae RNA Pol II general transcription factors and coactivators RNA polümeraas II struktuur
Rpb1 CTD (C-terminal domain) CTD on erakordselt tähtis RNA polümeraasi aktiivsuse regulatsioonil. CTD asub Rpb1 valgu C-terminuses ja koosneb 26-52 ühetaolisest kordusjärjestusest: YSPTSPS. Korduste arv on erinevatel organismidel erinev: pärm 26, C. elegans 32, Drosophila 45, imetajad 52. Kui Rpb1 küljest CTD ära võtta, siis selline polümeraas pole võimeline transkriptsiooni teostama. Samuti on vajalik, et vähemalt 2 kordust tuleksid järjest: kui CTD-le lisada aminohappeid nii, et need tuleksid iga korduse järel, siis selline polümeraas ei tööta, kui selliseid lisamisi teha iga kahe korduse tagant, siis töötab. Kui aga transkriptsiooni reaktsioon teostada puhastatud valkudega in vitro, siis ilma CTD-ta polümeraas siiski suudab RNA-d sünteesida, mis näitab, et CTD pole oluline otseselt RNA sünteesi reaktsiooniks, vaid tal on regulatoorne tähtsus. Nii seriini- kui ka treoniinijääke fosforüleeritakse, neist kõige olulisemateks peetakse Ser2 ja Ser5 fosforüleerimist. Väga üldises mõistes eristatakse RNA polümeraasil fosforüleeritud ja defosforüleeritud vormi (mis tegelikult tähendab CTD fosforüleerituse olekut). Fosforüleeritud polümeraas on transkriptsiooniliselt aktiivne, defosforüleeritud vorm on nn “vaba”: see mis on rakus saadaval, kuid ei ole parasjagu transkriptsiooni teostamas.
Rpb1 CTD (C-terminal domain) CTD fosforüleeritakse PIC kompleksis TFIIH poolt, kui polümeraas transkripsiooni lõpetab ja DNA-lt vabaneb, siis toimub CTD defosforüleerimine ning selline polümeraas on valmis uuesti PIC-iga seonduma. Tegelikult toimub transkriptsiooni käigus CTD fosforüleerimismustri muutus: promootoriga seondunud ja transkriptsiooni alustav RNA polümeraas on fosforüleeritud eelkõige Ser5 positsioonis, elongatsiooni käigus see modifikatsioon eemaldatakse ning asendatakse Ser2 fosforüleerimisega. Selle mustri selgitamisel oli küll veidi tehnilisi probleeme, mistõttu pole selge, kas elongatsiooni käigus on tegu ainult Ser2 või Ser2Ser5 fosforüleerimisega.
Rpb1 CTD (C-terminal domain) • Milleks CTD ja selle fosforüleerimine ikkagi vajalik on? • CTD fosforüleerimine on vajalik PIC-i küljest vabanemiseks. • CTD asub polümeraasi kompleksi selles piirkonnas kust väljub sünteesitav mRNA. CTD kaudu seonduvad polümeraasiga kõik mRNA protsessinguks vajalikud ensüümide kompleksid: CAP-struktuuri sünteesil osalevad valgud, splaissinguks vajalikud faktorid, transkripti lõikamiseks ja polyA sünteesiks vajalikud faktorid. • Sinna seonduvad kromatiini modifitseerivad ensüümid, mis on vajalikud polümeraasi teele jäävate nukleosoomide osaliseks või täielikuks lagundamiseks. • Sinna seonduvad ka CTD fosfataasid, mis transkriptsiooni lõppedes viivad polümeraasi tagasi defosforüleeritud olekusse. CTD defosforüleeritakse ka siis, kui polümeraas peaks elongatsiooni ajal liiga kauaks seisma jääma, näiteks DNA kahjustuse korral.