Download

1 / 17
310 likes | 564 Views
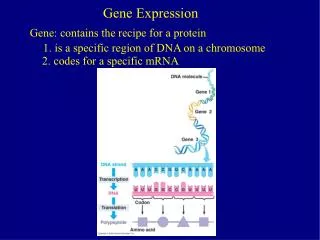
mRNA decay - regulating gene expression -. Wiebke Ginter 06.12.10. Differences of eukaryotic and bacterial mRNA. Bacterial mRNA - Triphosphate Stem-loop Ribosome binding: base pairing between the 3’ end of 16S ribosomal RNA and a Shine–Dalgarno element. Eukaryotic mRNA

E N D
mRNA decay- regulating gene expression - Wiebke Ginter06.12.10
Differences of eukaryotic and bacterial mRNA Bacterial mRNA - Triphosphate • Stem-loop • Ribosome binding: base pairing between the 3’ end of 16S ribosomal • RNA and a Shine–Dalgarno element Eukaryotic mRNA • 5’ 7-methylguanosine cap • 3’ poly(A) tail with poly(A)-binding protein (PABP) • Ribosome binding: affinity of the small ribosomal subunit for eukaryotic initiation factor 3 (eIF3)
Conventional pathways for mRNA degradation (E. coli) • - serial internal cleavage by RNase E • lack base pairing at the 3’ end • susceptible to attack by the 3’ exonucleases polynucleotide phosphorylase (PNPase), RNase II, RNase R and oligoribonuclease
Conventional pathways for mRNA degradation (Eukaryotes) • PAN2-PAN3 • PABP-dependent poly-A nuclease, 60-80nt • CCR4-NOT • 9 protein • exonuclease domains in Ccr4 and Caf1 • activity inhibited by PABP • PARN • Cap-dependent deadenylase • processivity enhanced by 5’cap • inhibited by cap-binding proteins • mass deadenalytion in maternal mRNA in oocytes (Xenopus), in various cell lines, embryogenesis in plants • Dcp1/2 • Decapping enzyme • dimer • XRN1 • exoribonuclease • degrades 5′→3′ direction • Exosome • Largecomplex of 3′→5′ exonucleases • 10-12 SU with RNase PH domain • homologies with hydrolytic exonucleases, RNA helicases
P-bodies Lsm1XRN1DNA • - Cellular sites of decay, but also RNA storage • - Granular cytoplasmic foci • Enriched in components of 5’ → 3’ decay • assemble when 5’ → 3’ decay system is overloaded with mRNA or decay is impaired
Unusual routes to decay • Deadenylation-independent decapping • bypass deadenylation step – directly decapped • autoregulatory • Rps28B directly binds stem-loop of 3’ UTR of own mRNA • recruits Edc3 – enhancer of decapping • association of other decapping factors Edc1: decapping regulator intramolecular pairing blocks access to the deadenylase: interaction between the poly(A) tail and a poly(U) stretch in the 3′ UTR feedback regulation
Unusual routes to decay • Endoribonucleolytic decay • PMR: polysome-associated endonuclease • Targeting actively translating mRNA • IRE1: endonuclease on endoplasmic reticulum • Targeting actively translating mRNA • MRP: multicomponent complex, RNase • Processing rRNA/nucleolus, mitochondrial RNA • In temporal asymmetric MRP bodies during mitosis
Non-sense mediated decay (NMD) - I • Detects premature termination codons (PTC) • arise from mutations, frame-shifts, inefficient processing, leaky translation initiation and extended 3’ UTR • truncated proteins with aberrant functions • Core proteins of the NMD complex: UPF1, UPF2 and UPF3 • exon junction complex (EJC) • feature of an aberrant transcript, residual ‘mark’ of splicing • 20–24 nucleotides upstream EJ • Also role in regulating normal gene expression
Non-sense mediated decay (NMD) - II • Detects premature termination codons (PTC) • arise from mutations, frame-shifts, inefficient processing, leaky translation initiation and extended 3’ UTR • truncated proteins with aberrant functions • Core proteins of the NMD complex: UPF1, UPF2 and UPF3 • exon junction complex (EJC) • feature of an aberrant transcript, residual ‘mark’ of splicing • 20–24 nucleotides upstream of every • Also role in regulating normal gene expression Most: deadenylation-independent decapping in P bodies
Non-stop decay • Targets mRNAs that lack a stop codon • Premature polyadenylation • facilitates the release of the ribosome • Ski-complex (Ski1,3,8) • Ski7 (adaptor) binds to empty A site • release ribosome • Ski7 recruits exosome • SKI-complex deadenylates • decay 3’→5’ direction • No Ski7: 5’ →3’ decay pathway (due to PABP removal)
No-go decay • Detecting stalled ribosomes • Endonucleolytically cleaving the mRNA • Dom34-Hbs1 needed for initial cleavage • decayed by the exosome and Xrn1
Signals that control mRNA decay • Modulation of RNA-binding proteins • mRNA=unstable=facilitate rapid changes if mRNP is changed • P38 MAPK, ERK, JNK, Wnt/β-catenin pathways influence ARE-function • Modulate mRNP structure, mediate phosphorylation of ARE-binding proteins, alter affinity, bind other factors • Puf proteins • Recognise UG-rich sequences • Accelerates decay • CCR4-NOT deadenylase recruited • Each Puf has special target transcripts • Regulate certain cellular processes • Stabilising elements • Sequence elements can confer stability = transcripts of housekeeping proteins = stable • Pyrimidine-rich elements in 3’ UTR • αCP1 and αCP2 bind • Protecting poly(A) tail from deadenylation • AU-rich elements (ARE) • Stability element • 3’UTR of cytokines, proto-oncogenes, transcription factors • AUUUA-pentamer – several classes • No 2 identical • Flanking region can influence overall effect on mRNA stability • Enhance decay by recruiting mRNA-decay machinery • Interacts with exosome (AUF1, TTP) • Bind PARN deadenylases (KSRP, RHAU) • Stabilising mRNA-binding proteins • Removing mRNA from decay sites? • Competing with binding sites for decay factors? • Inhibit decay machinery? • Strenghten PABP-poly(A) interaction?
Interfacing with other cellular mechanisms • Translation • General inhibitions of translation elongation → stabilising mRNAs on polysomes • Inhibtition of translation initiation → diverts transcripts to P-bodies for decay • Many mRNA-binding proteins that influence mRNA turnover also regulate translation • Transcription • CCR4–NOT complex represses RNA polymerase II required for both transcription and deadenylation • Rpb4 protein • subunit of RNA polymerase II • also required for deadenylation and decay • localizes to P bodies • essential role in modulating gene expression in response to stresses such as glucose deprivation and heat shock • mRNA localisation • DCP1 and CCR4 – implicated in localisation of mRNA transcripts
Bacterial decay Transient addition of poly(A) tails=crucial for exonucleolytic degradation of stem-loop structures Pyrophosphohydrolase: conversion of 5’ terminal triphosphate to monophosphate → more susceptible to 5‘ monophosphate dependent RNase=endonuclease RNase E Quality control: PTC Non-stop decay: tmRNA No-go decay: endonuclease Eukaryotic decay Poly(A) tail Resemblance to decapping (catalyses by related enzymes)=removing a protective group → more susceptible to 5‘ monophosphate dependent RNase=exonuclease XRN1 Quality control: PTC, recognise abnormal 3’ UTR Non-stop decay: Ski7 No-go decay: endonuclease Key differences mRNA decay
Bacterial decay Mostly by low specificity endonucleases Poor ribosome binding → decay (spacing increases, cleavage sites free) Shorter intercistronic and 3’ UTR Poly(A)= destabilising Internal ribosome binding sites – co-transcribed polycistronic operons possible – can also degrade discrete segments only Eukaryotic decay 3’ and 5’ terminal events=dominant (deadenylation, decapping, exosomes) Endonucleases=much less, more specific Inefficient initiation: not doomed to degradation 3’ UTR long, contains binding sites for regulating proteins Depend on deadenylation of stabilising poly(A)– need of protective PABP eIF4F protein complex governs terminal ribosome binding, interaction with PABP and poly(A) tail interupted by deadenylation Key differences mRNA decay