Download

1 / 1
10 likes | 134 Views
Survey of autofluorescence in the wings of dragonflies Pachydiplax longipennis and Erythemis simplicicollis with emphasis on costal morphology as related to flight mechanics Razin Ahmed and David Neff Department of Bio. Science, Marshall University. Stigma. Nodus. Subcosta. Costa.

E N D
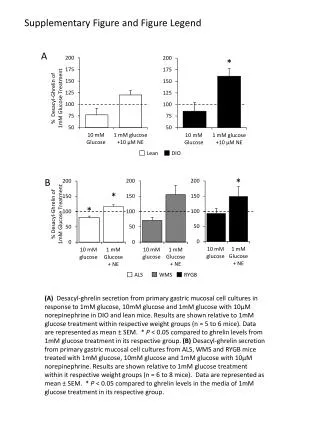
Survey of autofluorescence in the wings of dragonflies Pachydiplax longipennis and Erythemis simplicicollis with emphasis on costal morphology as related to flight mechanics Razin Ahmed and David Neff Department of Bio. Science, Marshall University Stigma Nodus Subcosta Costa Forewing 100 um Hind wing E. simp. triangle costal side view P. long. triangle costal side Fig. 6 E. simp. stigma side P. long. stigma side 1mm E. simp. stigma z-step 3D E. simp. stigma ventral z-step 3D Triangle 1mm Pachydiplax Erythemis Fig. 4 1 mm Pterostigma 1 mm Pachydiplax Fig. 7 Erythemis Fig. 1 ABSTRACT This study examined morphological variation in two species of dragonflies from the family Libellulidae, Erythemis simplicicollis (pond hawk) and Pachydiplax longipennis (blue dasher) [Fig. 1,2]. A review of the literature on the two species demonstrated substantial behavioral variations. Notably, male intraspecific territorial behavior has been described and consist of unique flight patterns. Classical dragonfly taxonomy is often based upon the venation of the wings which have been shown to be specialized to fulfill aerodynamic needs. The goal of this study was to see if the morphology of the wing structures of the dragonflies made any difference when it came to these unique flight patterns. To this end we will use a confocal laser scanning microscope to observe the autofluorescence that occurs in most insect cuticles; this data will be used to reconstruct the structures of interest. Particular features were chosen based on flight studies that have shown that variation in morphology of wing features can correlate with various biomechanically defined flight behaviors and patterns. Thus the morphologies of two distinct areas of wing venations and thickness of the wing blade, features common to all Libellulinae, are examined (Fig. 6). Also, ratios of whole wing dimensions and body mass are measured as a means to normalize the wing vein data. Fig. 5 A. B. Fig. 3 Introduction Dragonflies are highly dependent on their wings to perform their daily activities. Without their wings they would not be able to forage for food, mate, and defend their territory. Just as our ancestors gained inspiration from flying creatures in their quest for flight, so might we use the natural forms shaped by eons of selective pressure to inspire new ideas pertaining to flight and engineering. Dragonflies are better to study as aero models than for example, flies, because first they have the most unusual of insects flight behaviors, the ability to glide while not flapping wings. Secondly the way they fly and maneuver cannot be duplicated by any other order. Thirdly, as gliders, their wing structure (as an aerofoil) tells us much of why they are such great fliers. Lastly they are one of the largest insects so it can be easier to study them, especially in experiments done by researchers that employ simultaneous video/aerodynamic data collection. These qualities make them good models for fixed wing studies. Unlike aircraft, most winged insects must produce aerodynamic forces by flapping their wings, this is not always so with the dragonfly. “An aircraft has a pair of fixed wings, and the forces necessary for flight are generated by the interaction between the wing and the surrounding air” (Sudo et al.1999: pg.723). Sudo goes on to say that the gross weight in insects is equivalent to the lift generated during the hovering of the insect. For dragonflies, the dimensions of their wings let them be extremely versatile fliers. They are able to fly at high-speed forward, hovering, and backward. They can also fly by vertical takeoff and landing. However of all these abilities it is the dragonfly’s gliding that makes them useful in studying fixed aerofoils. According to Sudo and colleagues, leading edge morphology, camber, vein bifurcation angles, body weight, and gross dimensions of dragonfly wings all correlate to the flying behavior of dragonflies. The aerofoil is defined by 3 basic measurements, span, chord and camber. These qualities of a wing combine to define its glide ratio or coefficient of lift/coefficient of drag (Fig. 3, Fig. 5 A.). Dragonfly span and chord is the same measurement as in aircraft wings (Fig.4). Camber in dragonfly is defined by leading edge shape and by corrugations in an otherwise flat membranous wing. Among insects, only locusts have such severe corrugations and are such good gliders as well (Okamoto et al.). Wing of dragonfly like virtually all insect cuticle is made of chitin (sugar) fibers combined with various protein and non-protein compounds that can serve as cross-links. These fibers and cross-links define the material properties of the insect’s exoskeleton (including wings). Chitin itself doesn’t fluoresce brightly, however, the other components of cuticle have been shown to autofluoresce (Zill, 2000); here we use this property to create a representation of the dragonfly wing. The basic features of the wings are cells demarcated by veins and spanned by membranes. In the present study we have surveyed the wings of twodragonfly species, Pachydiplax longipennis and Erythemis simplicicollis. We have collected data that show the different regions of the wing emit qualitatively different light wavelengths. The species Pachydiplax longipennis and Erythemis simplicicollis were studied due to the fact that they were readily available around Huntington, WV and dry samples were available in the Biological Department of Marshall University. The two species from the same suborder (Anisoptera) and family (Libellulidae) differed in wing structure and flying behavior. The purpose of the study is to test for correlations of flying behavior to wing morphologies of Pachydiplax longipennis and Erythemis simplicicollis and to make a simple survey of fluorescent structures in the dragonfly wing. Conclusion and Discussion From the data collected and analysis of the results, it is plain that Erythemis simplicicollis is larger in size compared to Pachydiplax longipennis. Also the wings of the two species vary in several ways. The general shape of the leading edge (costal margin) is different in the two species, this can be confirmed by viewing the 3D reconstructions and side profiles that were compiled from serial optical sections. The 3D renderings of the costal margins show the leading edge of both species to be much thicker than the more caudal wing surfaces. Both are notably sharp which has been shown to contribute to a positive lift to drag ratio (Okamoto, 1995). The most notable inter-specific difference is the orientation of the leading edge. Also, while the aspect ratio (span/chord) in the two species is approximately the same (3.2:1), the wing area to mass ratio is considerably higher in Erythemis simplicicollis (15:1) when compared to Pachydiplax longipennis (10:1). Finally, Okamoto and his colleges said that the thinner wings have a better performance with a smaller drag coefficient and a larger maximum lift coefficient. They go on to say that the configuration at the leading edge seems to be important, the aerofoil with the downward-facing leading edge has a much better performance than the upward-facing model showing considerably better glide performance than a downward facing model ( Fig. 5 A.) [Okamoto et al., p. 287]. Antonia Kesel adds to this by saying that during gliding, dragonfly wings can be interpreted as acting as ultra-light aerofoils which, for static reasons, have a well-defined cross-sectional corrugation (Fig. 4 ). Furthermore, the orientation of the leading edge changes at the nodus, which is the first bend formed by the costa, subcosta and radius. This orientation changes because the costa and subcosta fuse at the nodus, distal from which, the first valley is built by the costa, radius and mediana 1, so that the leading edges faces downwards. Additionally, the fluorescent survey does indeed demonstrate spectral differences in the emissions coming from the veins as compared to light emitted from the membranes. The purple appearing areas map emitted light of wavelengths 580-700 nm. The areas appearing green/yellow map shorter wavelengths (490-610nm). Those whitish regions represent broad spectrum light. Study of the pterostigma yielded an interesting finding . In P. longipennis. the stigma fluoresced intensely and broadly while in E. simplicicollis stigma emissions were notably feeble. Also P. longipennis showed somewhat greater emission intensity across the spectrum from all areas. Our data that shows the different regions of the wing shows qualitatively different emission spectra raises the future possibility of correlating well defined mechanical properties with endogenous emissions. From the data gathered and the literature reviewed we conclude that the mapping of autofluorescent structures in insect wings can be a valuable tool in the analysis of wing biomechanics. Knowing the evolutionary background of these insects and understanding the relationship between structures that have autofluorescence and how these structures help the insect in forging for food, mating, and defending their territory, provide important insight to aid our understanding of animal flight mechanisms. This relatively unexplored field should provide opportunities for further inquiry and research. Pachydiplax C. Figure 5 A. is Okamoto’s data showing correlation between wing corrugations, leading edge shape, and lift:drag ratio (gliding performance). In B. and C. our 3D representations show that P. longipennis has a clearly downward facing angle to the leading edge while E. simplicicollis has a more symmetrical arrangement of the costal margin with respect to the main plane of the wing. Figure 4 shows an histological transverse section through a dragonfly wing (Okamoto). Note the corrugations that give the otherwise flat wing an aerofoil shape. The blue circle shows the location of our 3D reconstructions as seen in Fig. 5. Fig. 2 A. Triangle Materials and Methods Using past biomechanical (wind tunnel) studies as a guide, we chose to focus our search on the costal margin (Fig. 6). We also surveyed at low-mag. specific, chord-wise regions at mechanically important areas of the wings (triangle and pterostigma). Most data was collected with a laser scanning confocal microscope (Bio-Rad MRC 1024). Using this microscope and Bio-Rad software we were able to take optical sections of the wings at three different emission wavelengths and reassemble these optical sections into 3D renderings. Pachydiplax longipennis and Erythemis simplicicollis wings were cut up into three pieces and put into phosphate-buffer saline (PBS) solutions. The wings of the two species of dragonflies that were available were measured by a digital caliper and were weighed by a digital balance. We rehydrated dried specimens in PBS (137mM of NaCl, 2mM of KH2PO4, 2.7mM of KCl, 10mM Na2HPO4; pH tested were 2, 7, and 12). An initial survey of pH dependent fluorescence showed that PBS of pH 12 gave the highest fluorescent yield from the veins. We were pleased to find that old, dry specimens fluoresced adequately after rehydration. After finding optimal pH, left wings of both species were rehydrated in the buffer solution, cut chord-wise and placed between two coverslips. The areas defined above indicate where we cut the wings. These were then imaged under the confocal microscope and analyzed by 3D rendering and resampleing software. The laser band used for excitation was 488nm (blue). We collected only short red and long red (580-700nm) for leading edge studies. We collected three channels red (580-620nm), green (506-538nm) and long red (660-700nm) for whole wing survey. For leading edge studies, we collected a series of confocal images in the z (vertical) axis. Using Confocal Assistant and Lasersharp software, we reassembled these optical serial sections into a 3D representation of the wing veins and membranes. These renderings were resampled in the z axis; thus, we were able to create virtual cross sections of the leading costal margin. We sampled the margins anterior to the triangle (pre-nodal) and at the pterostigma (ante-nodal). After all the data was collected the pictures and measurements were compared and contrasted to see if any significant differences could be seen between the two species’ wings and if any hypotheses can be made as to how these relate to their flight patterns. Figure 7 shows the three channel confocal fluorescent images of the wings of both species. In A. are montaged images of the wing taken chordwise. The costal margin is at the left, these are left wings as viewed from above (dorsal aspect). In B. are images taken across the wing from the pterostigma to the caudal (hind) margin of the wing. The purple appearing areas map emitted light of wavelengths 580-700 nm. The areas appearing green/yellow map shorter wavelengths (490-610nm). Those whitish regions represent broad spectrum light. Notice the pterostigma which differs significantly in the two species and the proximal (top) corner of the triangles which shows broad spectrum emissions.. LITERATURE CITED Kesel, Antonia. (2000). Aerodynamic characteristics of dragonfly wing sections compared with technical aerofoils. The Journal of Experimental Biology. 203, 3125-3135. Mitchell, Forrest. 1997. Digital Dragonfly Museum. http://stephenville.tamu.edu/~fmitchel/dragonfly/index.html Mullins, Lee Ann. 1994. Behavioral and Physiological Ecology of Dragonflies (Odonata: Anisoptera) at the Green Bottom wildlife Management Area.. A thesis presented to the faculty of the Biological Science department of Marshall University. Needham, James. 1954. A Manual of the Dragonflies of North America, including the Greater Antilles and the Provinces of the Mexican Border. University of California Press Berkeley and Los Angeles. Okamoto, M., Yasuda, K., and Azuma, A. (1995). Aerodynamic characteristics of the wings and body of a dragonfly. The Journal of experimental Biology. 199, 281-294. Silsby, Jill. 2001. Dragonflies of the world. Smithsonian Institution Press. Sudo, S., Tsuyuki, K., Ikohagi, T., Ohta, F., Shida, S., and Tani, J. (1999). A study on the wing structure and flapping behavior of a dragonfly. JSME International Journal. 42, 721-729. Zill, S. (2000) 3D graphic reconstruction of the insect exosleton through confocalimaging of endogenous fluorescence. Microsc Res Tech. 48 ( 6), 367-384. B. Erythemis Acknowledgments I would like to thank first and foremost Mr. David Neff for all his hard work and dedication for this research. Also thank you for showing and letting me use the high powered specialized machines. A special thanks goes out to Dr. Strait for all her help and guidance. Finally, thank you to Liz Fet and everyone else who helped in anyway to make this research a reality. The Morphometrics Lab (S307) was supported by a NSF-CCL1 grant to Drs. Strait, Norton, Ghosh.