Download

1 / 20
210 likes | 442 Views
LECTURE 18: RNA TRANSCRIPTION, PROCESSING, TURNOVER. Levels of specific messenger RNAs can differ in different types of cells and at different times in the same cell. Control of RNA abundance can be at the level of transcription initiation, transcription elongation, processing or degradation.

E N D
LECTURE 18: RNA TRANSCRIPTION, PROCESSING, TURNOVER Levels of specific messenger RNAs can differ in different types of cells and at different times in the same cell. Control of RNA abundance can be at the level of transcription initiation, transcription elongation, processing or degradation. RNA control mechanisms are more complex and varied in eukaryotes than in prokaryotes.
RNA POLYMERASE Mediates transcription of RNA from a double-stranded DNA template Does not require primer Transcription is always 5’ --> 3’ direction, with each base addition using reaction RNAn + NTP --> RNAn+1 + PPi Transcription inititation mediated by proteins that bind to specific promoter sequence elements. The promoter binding proteins recruit RNA polymerase to the initiation site. RNA polymerases show evolutionary conservation between prokaryotes and eukaryotes, accounting for many similarities in transcription mechanism But there are substantial differences in transcription initiation, termination, and post-transcriptional RNA processing between pro- and eukaryotes
PROKARYOTIC TRANSCRIPTION INITIATION Prokaryotic promoters have elements near position -10 and -35 that bind to the s initiation factor. Promoter elements have been physically mapped by DNA footprinting. Footprinting Technique
TRANSCRIPTION - TRANSLATION COUPLING IN PROKARYOTES Prokaryotic mRNA does not require processing nor cell compartment trafficking to become competent for translation. Therefore, transcription and translation are coupled. In prokaryotes, gene regulation is almost exclusively at level of transcription initiation, and not later transcription or translation processes.
PROKARYOTIC TRANSCRIPTION TERMINATES BY RNA MECHANISM r (rho) - independent transcription termination signal motif r protein is recruited to RNA polymerase during elongation to allow transcription termination at other signal motifs
PROKARYOTIC RIBOSOMAL RNA IS GENERATED BY ENDONUCLEASE PROCESSING OF A PRECURSOR TRANSCRIPT
THREE EUKARYOTIC RNA POLYMERASES SYNTHESIZE DIFFERENT TYPES OF RNA BY DIFFERENT INITIATION MECHANISMS
POL II INITIATION COORDINATED THROUGH A TATA-BOX PROMOTER Transcription initiation factors were painstakingly identified through establishment of cell-free in vitro transcription assays using TATA-box- containing DNA fragment, RNA Pol II, 32P-NTPs, and nuclear protein extracts. Competent extracts were then subjected to biochemical fractionations and reconstitutions.
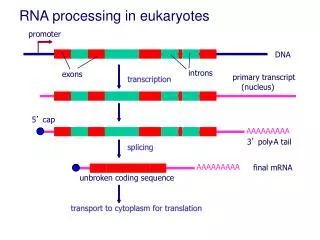
EUKARYOTIC POST- AND CO-TRANSCRIPTIONAL RNA PROCESSING Transcription elongation very similar in eukaryotes and prokaryotes: moving polymerase transcription bubble. Little is known about eukaryotic transcription termination. 3’ ends of Pol II transcripts are generated by site-specific endonuclease cleavage and template-independent poly-adenylation. s
EUKARYOTIC POST- AND CO-TRANSCRIPTIONAL RNA PROCESSING Nascent Pol II transcripts undergo capping of the 5’ end through reaction with GTP in a 5’-5’ orientation. Capping protects RNA from exonuclease degradation and provides a recognition site for translation initiation factors after export to the cytoplasm.
EUKARYOTIC POST- AND CO-TRANSCRIPTIONAL RNA PROCESSING Nascent Pol II transcripts in most cases undergo splicing to remove intronic sequences and unite coding sequence exons.
POL II TRANSCRIPTION AND PROCESSING ARE COUPLED IN NUCLEUS Capping, splicing, and 3’ cleavage/polyadenylation all occur while RNA is being transcribed.
EXON CHOICE IN SPLICING Most frequently, splicing joins adjacent exons to assemble mature RNA. For some genes, exons are “skipped” during splicing to generate alternative RNA products. Exon “choice” is tightly regulated, allowing generation of different proteins from same gene in different cells. Different growth factor (FGF) receptors are synthesized due to selective use of exons 5, 6, or 7 into the mRNA. Epithelial cells use exon 6 (IIIb), but mesenchymal cells use exon 7 (IIIc). The alternative receptors bind to different FGFs.
IMPROPERLY SPLICED RNAs ARE DEGRADED IN THE CYTOPLASM BY NONSENSE-MEDIATED mRNA DECAY Not all transcripts being made from a gene get spliced correctly. Many mis-spliced RNAs will contain nonsense codons and generate a highly truncated protein. Mis-splicing frequency can be enhanced by mutation. WHY and HOW is the abnormal mRNA degraded???
IMPROPERLY SPLICED RNAs ARE DEGRADED IN THE CYTOPLASM BY NONSENSE-MEDIATED mRNA DECAY As a messenger RNA is first translated, EJC proteins are removed as the ribosome passes by. Normally, translation termination will occur after all EJCs have been deactivated. If there is premature termination, the translation release factor can interact with remaining EJC proteins, providing signal for degradation.
FOR SOME GENES, NONSENSE MEDIATED mRNA DECAY IS USED TO AUTOREGULATE LEVEL OF GENE EXPRESSION Pyrimidine tract binding (PTB) protein feeds back to induce mis-splicing of PTB pre-mRNA, thereby causing its cytoplasmic degradation by nonsense method.
MEASURING RATES OF A SPECIFIC RNA’S SYNTHESIS AND BREAKDOWN Northern blotting, RNAse protection assay, or RT-PCR are used to measure amount of a specific RNA exists in a particular cell or tissue at particular time. Differences in a gene’s RNA levels detected by these assays DO NOT reveal what causes the RNA levels differ. Nuclear run-on transcription assay measures ongoing rate of a gene’s transcription at a particular region along the gene. Therefore, this assay can determine whether differences in a genes total RNA abundance is the result of different rate of synthesis OR different degree of completion, as some genes have “transcription attenuation” sites. Inhibition of new RNA synthesis with toxins (e.g. Actinomycin D) can be used to measure the stability (half-life) of a mature RNA species. Or drive transcription of specific RNA ectopically from strong promoter and measure steady-state abundance.
EXAMPLES OF RNA CONTROL MECHANISMS Growth factors and hormones control rates of transcription initiation at specific genes (nuclear run-on assay) C-myc proto-oncogene expression level is controlled by transcription attenuation. Some cancer cells show failure to induce c-myc transcription attenuation Heat shock controls levels of Hsp70 RNA by suppressing a rapid RNA degradation mechanism



















![[V]. Process of Transcription and Transcriptional Control of Gene Expression](https://cdn2.slideserve.com/5058527/v-process-of-transcription-and-transcriptional-control-of-gene-expression-dt.jpg)