Download

1 / 44
480 likes | 958 Views
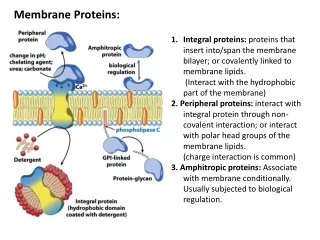
Types of membrane proteins. Transmembrane proteins Cytosolic membrane proteins External membrane proteins Peripheral membrane proteins. Types of membrane proteins. 4. Peripheral membrane proteins

E N D
Types of membrane proteins • Transmembrane proteins • Cytosolic membrane proteins • External membrane proteins • Peripheral membrane proteins
Types of membrane proteins 4. Peripheral membrane proteins • Proteins on either side of the membrane that associate with the polar ends of transmembrane proteins by noncovalent (ionic) interaction (Fig. 10-17)
Peripheral Membrane Proteins Noncovalent association with the polar ends of transmembrane proteins
How do transmembrane proteins cross the bilayer? • Most do so in an α-helix conformation • α -helix composed largely of hydrophobic amino acids that interact with the hydrophobic fatty acids of phospholipid (Fig. 10-19)
Hydrophobic amino acids in light green & yellow
Glycophorin Single-pass transmembrane protein with an α helix transmembrane domain
How do transmembrane proteins cross the bilayer? • Most do so in an α-helix conformation • α -helix composed largely of hydrophobic amino acids that interact with the hydrophobic fatty acids of phospholipid • Peptide bonds are polar • Because of absence of water in an α -helix , peptide bonds driven to form hydrogen bonds with each other. (Fig. 3-9)
Ribbon + C and N backbone Arrangement of atoms showing H bonds Ribbon illustration of α-helix The N-H of every peptide bond is hydrogen-bonded to the carbonyl group of the peptide bond 4 amino acids away
How do transmembrane proteins cross the bilayer? 2. Other proteins traverse the bilayer as an antiparallel β sheet, forming a β barrel • This also allows peptide bonds in a lipid bilayer to hydrogen bond (Fig. 3-9)
Antiparallel β sheet (peptide chains run in opposite directions) Peptide chains held together by hydrogen bonds between peptide bonds of adjacent peptide chains
β-barrel proteins • Some β barrels form transmembrane channels for transport • Very abundant in the outer membrane of mitochondria and chloroplasts & bacteria • They form water-filled pores • e.g. the porin barrel • Formed from a 16 stranded antiparallel β sheet • Rolls up into a cylindrical structure (Fig 10-21)
Polar amino acid residues line the aqueous channel on the inside Nonpolar amino acid residues project from the outside and interact with the lipid fatty acid sidechains
Hydropathy plots • Membrane proteins are difficult to crystallize because of their hydrophobicity • Accordingly, very few have been analyzed by X-ray crystallography, the standard technique to resolve molecular structure.
Hydropathy plots • However, DNA cloning and sequencing have enabled identification of amino acids of membrane proteins • Structural information can be inferred from this sequence information • Specifically, can predict where in the protein transmembrane segments reside from a determination of hydrophilic and hydrophobic amino acids • Transmembrane segment is typically 20-30 amino acids with high degree of hydrophobicity (Fig 10-20)
1 membrane-spanning α helix Hydrophobic 7 membrane-spanning α helices Hydrophilic Hydropathy Plots
Solubilization of membrane proteins • Transmembrane proteins are difficult to solubilize because of their hydrophobicity • Requires detergents • Small amphipathic molecules that form micelles in water (Fig. 10-23)
Cross section of a detergent micelle SOLUBILIZATION The hydrophobic ends of the detergent bind to the hydrophobic region of the transmembrane protein, thereby displacing lipid molecules Since the other end of the detergent molecule is polar, the protein is brought into aqueous solution as a detergent-protein complex
Detergent solubilization of membrane proteins
Solubilization of membrane proteins • Transmembrane proteins are difficult to solubilize because of their hydrophobicity • Requires detergents • Small amphipathic molecules that form micelles in water (Fig. 10-23) • By using detergents, transmembrane proteins can be solubilized and reconstituted into liposomes • Powerful means of analyzing their properties (Fig. 10-26)
Na+- K+ pump solubilized & reconstituted into liposome (ion pump present on plasma membrane of most animal cells – uses ATP for energy) Pumps Na+ out of the cell and K+ into the cell
Sidedness of the RedCell Membrane • Easiest plasma membrane to isolate – hence well studied • Reason: • No internal organelles; • Hence, plasma membrane is the only membrane in red cell and can be easily isolated without concern about contamination by other types of membrane(Figs 10-27 & 28)
Red Cells are biconcave in shape and lack a nucleus and other organelles
Cytosol (mainly hemoglobin) is released Isolation of sealed and unsealed red blood cell Ghosts (plasma membrane) Can get right-side-out and inside-out vesicles depending on the ionic conditions of the mechanical disruption medium
Sidedness of the RedCell Membrane • Sidedness of proteins can be determined by vectorial labeling • use of a covalent reagent (radioactive or fluorescent) that binds to the portion of the protein exposed on the membrane surface • Reagent needs to be water-soluble so it won’t permeate the sealed ghost
Sideness of the RedCell Membrane • For example: • label sealed right-side out vesicles • Separate proteins by SDS-polyacrylamide gel electrophoresis (SDS-PAGE) • Labeled proteins detected by their radioactivity(autoradiography) or their fluorescence (expose to UV light) (Fig. 10-29)
SDS-PAGE separation of red blood cell proteins after labeling of sealed right-side-out vesicles A, gel stained with coomassie blue B, drawing of the positions of some major proteins Glycophorin labeled indicating it is exposed on the outer surface of the membrane
SDS-PAGE separation of red blood cell proteins after labeling of sealed INSIDE-out vesicles A, gel stained with coomassie blue B, drawing of the positions of some major proteins Glycophorin labeled indicating it is exposed on the INNER surface of the membrane as well Thus, the type of membrane proteins can be discerned
Diagramatic illustration of glycophorin in the Red blood cell membrane Transmembrane protein Exposed end on the outside surface Exposed end on the inner cytosolic surface
Major plasma membrane proteins of the Red Blood Cell • By SDS-PAGE, has been established that there are 6 major proteins • There are many more as well, but these are the main ones (i.e. present in highest concentration)
Major plasma membrane proteins of the Red Blood Cell • Spectrin (peripheral protein on cytosolic surface) • Long thin flexible rod • Principal component of the cytoskelton (protein meshwork underlying surface of the red cell)
Major plasma membrane proteins of the Red Blood Cell • Spectrin (peripheral protein on cytosolic surface) • Maintains structural integrity of the red cell (e.g. biconvave shape) • Necessary as red cells go through small capillaries (Fig. 10-30)
Heterodimer of 2 antiparallel polypeptide chains, termed α and β Heterodimers self-associate, head- to- head, at the phosphorylated end to form tetramers Electron micrographs of isolated spectrin
Major plasma membrane proteins of the Red Blood Cell • Spectrin (peripheral protein on cytosolic surface) • Tetramers linked together (& to band 4.1 protein) by actin filaments
Major plasma membrane proteins of the Red Blood Cell • Spectrin (peripheral protein on cytosolic surface) • Tetramers linked together (& to band 4.1 protein) by actin filaments • This forms a junctional complex • Serves as deformable protein mesh, allowing red cell to withstand stress on its membrane as it is forced through small capillaries (Fig 10-31)