Download

1 / 16
160 likes | 272 Views
Discover the fascinating mechanics of bacterial chemotaxis as they navigate gradients in time, not space. Learn about swimming patterns, flagellar motors, and molecular receptors that drive directed motion in bacteria. 8

E N D
Bacterial Chemotaxis • Bacteria swim toward attractants and away from repellents. • Their motion is a biased random walk due to control of tumbling frequency. • They measure gradients in time, not space. They adapt over time. • Chemotaxis probably relies on clustering and cooperation among receptors. Jason Kahn, UMCP
Fundamental mechanics of chemotaxis • Textbooks and Review Sources: • Macnab, R. M. (1987). Motility and chemotaxis. In Escherichia coli and Salmonella typhimurium: Cellular and molecular biology. 1st ed. (Neidhardt, F. C., ed.), pp. 732-759. Am. Soc. Microbiol., Washington, D.C. • Stock, J. B. & Surette, M. G. (1996). Chemotaxis. In Escherichia coli and Salmonella typhimurium: Cellular and molecular biology. 2nd ed. (Neidhardt, F. C., ed.), pp. 1103-1129. Am. Soc. Microbiol., Washington, D.C. • White, D. (2000). The physiology and biochemistry of prokaryotes. 2nd ed., Oxford University Press, Oxford. • Parkinson, J. S. (2004). Signal amplification in bacterial chemotaxis through receptor teamwork. ASM News 70, 575-582. Jason Kahn, UMCP
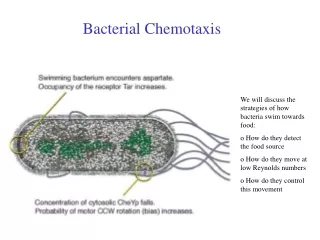
Swimming driven by a flagellar motor • The flagellum is an organelle consisting of (1) a basal body that is a rotary motor, (2) a connecting hook, and (3) a long filament. • Counter-clockwise rotation leads to all of the filaments of the cell twining together about a common axis, and swimming in a straight line. • Clockwise rotation leads to separation of separate filaments and tumbling, leading to random reorientation of the cell. • Typically the cell swims for 1 sec at 25 um/sec and then tumbles for 0.1 sec. A random walk. Chemoattractants increase the time spent swimming when the bacterium is going up the gradient. A biased random walk. Jason Kahn, UMCP
Bacterial Outboard Motor • The flagellum is a complex molecular motor driven by the transmembrane proton potential (pH) generated from oxidative phosphorylation or a proton-pumping ATPase. • Regulation and mechanism of assembly of flagellar subunits is a subject of current research. Jason Kahn, UMCP
Tumbling and the Biased Random Walk • From Macnab, 1987, E. coli and S. typhimurium • Left hand side shows CCW flagellar rotation. The corkscrewing action of the left-handed helical filament drives the bacterium in a straight line. Right hand side showsCW flagellar rotation, with the different filaments separating and driving in different directions, tumbling the cell. • Increasing the time spent swimming between tumbles leads to netmotion toward attractant. Decreasing the time spent swimming leadsto motion away from a repellent. Jason Kahn, UMCP
Assays for chemotaxis • Swarm plates: on semisolid agar, observe motion away from thecenter of a self-generated nutrient gradient. • Microscopic observation of free-swimming cells. • Tethering to an immobile surface allows observation of the rotation of the body of the cell. • Temporal stimulation (rapid changes in chemoeffector concentration). From Berg, Physics Today, 1999 Jason Kahn, UMCP
Dynamics of Chemotaxis • Bacteria respond to temporal, not spatial signals. Given their size, the spatial gradient is undetectable unless they move! • Thus they have a short term “memory” of the chemoeffector concentration over the previous few seconds. • A change in concentration leads (after a brief latency phase) to excitation (a change in tumbling frequency) followed by adaptation: after a few seconds at a new effector concentration the cell resumes random tumbling. This allows the cell to continue following a gradient up or down. Jason Kahn, UMCP
MCP Receptors and Stimuli • MCP = methyl-accepting chemotaxis protein. These are specifically engaged in chemotaxis, whereas periplasmic binding proteins for nutrients are primarily for transport. • Aer: Energy-linked attractants (O2). Monitors redox state of the cell. • Tar: Aspartate attraction, maltose attraction via binding MBP, the periplasmic binding protein (PBP) for maltose. Also Co2+ and Ni2+ repulsion • Tsr: Serine attraction, external pH • Trg: Sugar attraction via PBP’s • Tap: dipeptide attraction via PBP’s • Organic repellents act at much higher concentrations: may screw up membranes • Figure from Parkinson foreshadows higher-level organization Jason Kahn, UMCP
Che Proteins: CheA • Che = Chemotaxis genes. (Mot = motility, and many other flagellum mutants are defective in chemotaxis as well.) • Chemotaxis is a classic bacterial two-component signalling system. • Is general, these systems have a histidine kinase sensor protein and a response regulator that is controlled by phosphorylation via phosphotransfer from the kinase • The chemotaxis genes are CheA, CheW, CheY, CheZ, CheR, and CheB. • Extensively studied! • CheA histidine kinase is a dimer in solution. It autophosphorylates using phosphate from ATP: CheA + ATP CheA~P + ADP (on His 48) • CheA auto-phosphorylation is enhanced by CheW Jason Kahn, UMCP
Tar Structure and Interactions • Extracellular ligand-binding domain binds chemoeffector. • Signal is somehow transduced inside (through helix translation or rotation?) • Cytoplasmic helical domains interact with CheA, CheW • Ligand binding domain from 2LIG (Pymol), Stock 1997 Jason Kahn, UMCP
Tar-CheA-CheW ? • Cryo-EM image reconstruction / fitting X-ray structure model for the cytoplasmic domain of Tar interacting with CheA and CheW. (Francis et al., PNAS 2004). Suggests 6 receptor dimers per CheA dimer, plus 3 CheW monomers • Each red or yellow pillar is a trimer of dimers of TAR, but not the trimer of dimers seen in a crystal structure. • Controversial, mechanism of signalling to CheA unresolved. Jason Kahn, UMCP
Signal Transduction: CheY~P • Phosphorylated CheA transfers its phosphate to CheY, the response regulator: CheA~P + CheY CheA + CheY~P (on Asp 57) • CheY~P interacts with FliM and other flagellar proteins to switch the motor to CW rotation: tumbling. • Thus inactivation of CheA upon attractant binding to MCP’s leads to more swimming and less tumbling. Jason Kahn, UMCP
Stimulation: Control of CheY • CheY has an autophosphatase activity stimulated by CheZ: CheY~P + H2O CheY + Pi • So the steady-state level of CheY~P leads to the swimming / tumbling random walk, and the steady-state level changes in the presence of chemoeffectors. • CheZ may be unregulated, may interact with CheA. • This is a rapid response (< 1 sec) CheZ Jason Kahn, UMCP
Adaptation: Methylation • Adaptation means that bacteria stop responding to chemoeffectors when their concentrations do not change. They have a memory of the concentration over the previous few seconds. Hence adaptation is slower than excitation, allowing for a brief episode of biased random walking after a change in chemoeffector level. • Adaptation is mediated by the CheR methyltransferase and the CheB methylesterase. • CheR methylates the MCP’s using (Tar, Tsr, et al.). MCP-Glu + S-Adenosyl-Methionine (SAM) MCP-Glu-CH3 + Adenosyl-homocysteine • Methylated MCP’s are less responsive to attractant: CheA is not inactivated, CheY continues to be phosphorylated, bacteria continue to tumble. • There are 4 or 5 methylation sites per MCP, and if they occur in clusters this means there are many possible methylation states for a cluster of receptors: wide dynamic range. • CheR is slow, leading to the delay time in adaptation. Jason Kahn, UMCP
Adaptation II • CheB methylesterase removes methyl groups from MCP’s: MCP-Glu-CH3 + H2O MCP-Glu + CH3OH • Allows system to reset to be responsive to low concentrations of attractants again. • CheB is activated by phosphorylation by CheA. • Thus, CheA causes immediate excitation via phosphorylation of CheY, but also initiates a slower negative feedback loop that reduces the sensitivity of the MCP’s that regulate it! • MCP’s have allosteric responses to attractants that make them more susceptible to methylation: damping. • CheB auto-dephosphorylates slowly, perhaps unregulated. Jason Kahn, UMCP
Overall Summary of Circuitry Jason Kahn, UMCP