Download

1 / 28
390 likes | 1.14k Views
Chapter 6 Enzyme. 6.1 An Introduction to Enzyme. Highly specialized proteins with definitive functions Enzyme 1. Substrate specificity 2. Acceleration of specific chemical reaction 3. Mild conditions of temperature and pH. History of Enzyme Research. 1700s ~ 1800s

E N D
Chapter 6 Enzyme
6.1 An Introduction to Enzyme Highly specialized proteins with definitive functions Enzyme 1. Substrate specificity 2. Acceleration of specific chemical reaction 3. Mild conditions of temperature and pH
History of Enzyme Research • 1700s ~ 1800s • Recognition of biological catalysis • Digestion of meat by secretions of the stomach, conversion of starch to sugar by saliva • 1850’s • Vitalism • Louis Pasteur : alcohol fermentation by yeast is catalyzed by ‘ferments’ inseparable from the live cells • 1897 • Alcohol fermentation by yeast extract (Eduard Buchner) • Enzyme mediates the reaction (Frederick W. Kuhne) • 1926 • Isolation and crystallization of urease (James Sumner) • Enzymes are proteins
Most Enzymes are Proteins • Enzymes • Proteins >> RNA • Cofactor: chemical component of enzyme • Inorganic ions : Fe2+, Mg2+, Mn2+, Zn2+ …. • Coenzyme : complex organic or metalloorganic molecule • Transient carrier of specific functional group • Derivatives of vitamins • Apoenzyme + cofactor = holoenzyme
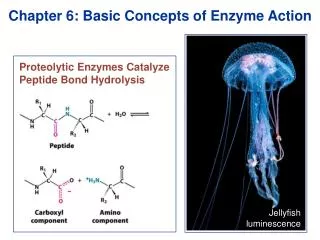
Classification of Enzymes • Classification by 6 catalytic functions • ATP + D-glucose ADP + D-glucose 6-phosphate • “ATP:glucosephosphotransferase” (hexokinase; common name) • Enzyme Commission number (E.C. number) : 2.7.1.1 • 2 : class number (transferase) • 7 : subclass (phosphotransferase) • 1 : hydroxyl group as acceptor • 1 : D-glucose as acceptor
Enzyme Affects Reaction Rate, Not Equilibrium • Reaction coordinate diagram • Free energy changes for reactions • DG’ 0 :biochemical standard free energy change (at pH 7.0) • DG‡ : activation energy • The difference between ground state and transition state energy levels • Energy barrier to chemical reaction • Catalysts enhance reaction rate by lowering activation energy
Enzymatic reaction • E + S ES EP E + P • Reaction intermediates • Any species on the reaction pathway that has a finite chemical lifetime (~ 10-13 sec : longer than molecular vibration) • e.g. ES, EP • Rate-limiting step • The step with the highest activation energy in a multi-step reaction
Thermodynamic definitions of equilibria and reaction rate • Reaction equilibrium • Equilibrium constant, Keq • K’eq =[P]/[S] • DG’ 0 = -RT lnK’eq • Reaction rate • Rate constant, k • V = k[S] : first-order reaction, k : s-1 • V = k[S1] [S2] : second-order reaction, k : M-1s-1 • From transition-state theory • k = (kT/h) e -DG‡ /RT • k :Boltzmann constant • H : Planck’s constant
Catalytic Power and Specificity of Enzymes • Enzymes as catalysts • Rate enhancement & High specificity • How enzyme works • Rearrangements of covalent bonds during an enzyme-catalyzed reaction • Transient covalent bond formation between catalytic functional groups of enzyme and a substrate • Transient group transfer to enzyme • Non-covalent interactions between enzyme and substrate • Binding energy DGB • The energy derived from enzyme-substrate interaction • Using DGB to lower activation energy • Generation of catalytic power using released free energy • Optimization of weak interactions in the reaction transition state
Weak Interactions Between Enzyme and Substrate • Enzyme must be complementary to the reaction transition state, not the substrate itself • Binding energy offsets the activation energy lower a net activation energy
Binding Energy Contributes to Reaction Specificity and Catalysis • Quantitative demonstration of binding E • DG‡ ; lowered by 5.7 kJ/mol increase k by10-fold • Energy available from a single weak interaction; 4 ~ 30 kJ/mol • Large enough to enhance enzyme reaction rate Enzyme’s reaction specificity & catalysis • Specificity • Ability to discriminate b/t a substrate and a competing molecule • Derived from the formation of many interactions between the enzyme and substrate
Enzyme catalytic mechanism • Binding energy to overcome barriers • Entropy reduction Restriction in the relative motions • Desolvation of the substrates Replacing water-substrate interactions to enzyme-substrate interactions • Thermodynamic compensation E involving weak interactions formed only in the transition state • Induced fit Proper alignment of catalytic functional groups (by conformational changes of enzyme) • Physical & thermodynamical barriers for DG‡ • Entropy (random of motion) • Solvation shell of water • Distortion of substrate • Need for proper alignment of catalytic functional groups
Enzyme catalytic mechanism Rate enhancement by entropy reduction
Specific catalytic groups contributes to catalysis 1. General Acid-Base Catalysis 2. Covalent Catalysis 3. Metal Ion Catalysis
General Acid-Base Catalysis • Unstable charged intermediate • Rapid break down to the reactants Inhibiting the reaction • Stabilized by protonation or deprotonation • Specific acid-base catalysis • Water as proton donor or acceptor • Faster proton transfer than break down of the intermediate no enzyme necessary • General acid-base catalysis • Other molecules as acid or base • Weak organic acids & base • Amino acid side chains in enzyme • The major catalytic mechanism in enzymes
Covalent Catalysis • Transient covalent bond formation between the enzyme and the substrate A-B A + B • With covalent catalyst (nucleophile X: ) • A-B + X: A-X + B A + X: + B • Lower activation energy than the uncatalyzed pathway • Nucleophiles • Amino acid side chains • Functional groups of enzyme cofactor H2O H2O
Metal Ion Catalysis • Ionic interactions between an enzyme-bound metal and a substrate • Orientation of the substrate for reaction • Stabilization of charged reaction transition states • Mediation of oxidation and reduction reactions • 1/3 of known enzymes require metal ions for catalytic activity
6.3 Enzyme Kinetics as an Approach to Understanding Mechanism
Enzyme Kinetics “Rate of reaction and changes of reaction rate in response to environmental parameters” • Initial rate (or initial velocity); Vo • Initial (< 60 sec) reaction rate when [S] is much higher than [E]
Enzyme Kinetics • Vo vs. [S] • Hyperbolic saturation kinetics • Formation of ES is a necessary step
General Theory of Enzyme Kinetics • Assumption for enzyme reaction • Fast reversible step • E + S ES • Slow rate limiting step • ES E + P • Saturation kinetics • At low [S] : Vo increases as [S] increases • At high [S] : Saturation of E with S • Approaching maximum velocity: Vmax • Steady state kinetics • Pre-steady state : Building up [ES], msec • Steady state : Approximately constant [ES] • Measured Vo state reflects the steady state k1 k-1 k2 k-2
Michaelis-Menten equation Chemical Equilibrium Michaelis-Menten postulation Steady State Approximation Steady state kinetics Pre-steady state kinetics rates of [ES] formation and breakdown
Michaelis Constant : Km • Michaelis-Menten equation • Vo =Vmax [S] / (Km + [S]) • When Vo = ½ Vmax • ½ = [S] / (Km + [S]) • Km = [S] • Km • [S] at Vo = ½ Vmax
Lineweaver-Burk Equation • Michaelis-Menten equation • Vo =Vmax [S] / (Km + [S]) • Lineweaver-Burk Equation • 1/Vo = (Km + [S]) / Vmax [S] • 1/Vo = Km / (Vmax [S]) + [S] / (Vmax [S]) • 1/Vo = Km / (Vmax [S]) + 1 / Vmax • Double reciprocal plot • 1/Vo vs. 1/[S] • Slope = Km / Vmax • Intercept on the 1/Vo axis = 1/ Vmax • Intercept on the 1/[S]axis = -1/ Km • Advantage of Double reciprocal plot • Accurate determination of Vmax • Distinguishing enzymatic reaction mechanisms • Analyzing enzyme inhibition