Download
1 / 19
190 likes | 345 Views
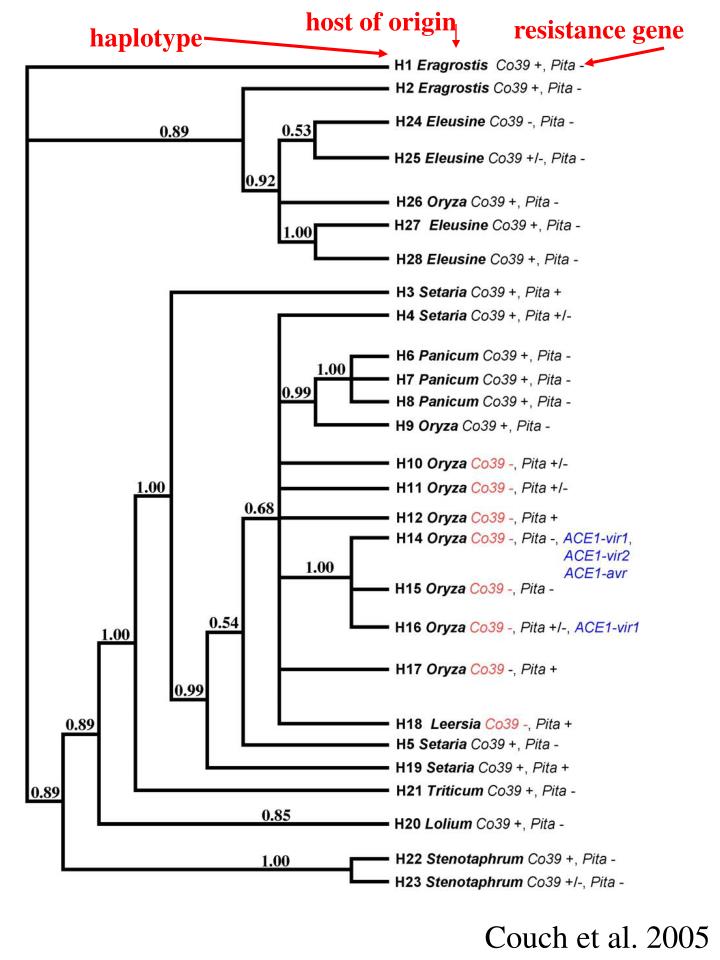
host of origin. resistance gene. haplotype. Couch et al. 2005. Magnaporthe oryzae. stage of. development signal. receptor. +. X. adenyl. cyclase. MAPK. transcription. factor. Signaling on the leaf surface. positive regulation. negative regulation. heterotrimeric G protein

E N D
host of origin resistance gene haplotype Couch et al. 2005
stage of development signal receptor + X adenyl cyclase MAPK transcription factor Signaling on the leaf surface positive regulation negative regulation heterotrimeric G protein dissociation MAPK protein kinase
spore tip mucilage Howard and Valent 1996
Cosegregation of the appressorium deficiency phenotype with the mpg1::Hph deletion allele. mpg1- MPG1 directs formation of the hydrophobic rodlet layer of conidia. WT hygS hygR mpg1- WT hygS hygR WT MPG1+ mpg1- MPG1+ mpg1-;hygR MPG1-;hygS Reintroduction of MPG1 restores pathogenicity, appressorium development and cell surface hydrophobicity
Importance of melanized appressoria Howard, R
albino WT chitin melanin appressorium pore Howard and Valent 1996
Penetration and invasion penetration site Dean et al, 2005
melanin biosynthesis Howard and Valent 1996
conidium germ tube appressorium Collapse high concentration of non-permeable solute sonicated cells Howard and Valent 1996
Appressoria build turgor during incubation Howard et al., 1991
The transcription factor MST1 is important for penetration peg formation homeodomain Zn finger WT mutants Park et al. 2004
can’t penetrate no peg The transcription factor MST1 is important for penetration peg formation mst1- WT appressoria can form From Park et al 2004
AVR-Pi-ta Fungal avirulence genes and others Lauge and de Wit 1998 four Avr genes; Avr2, Avr4, Avr4E and Avr9 four extracellular protein (Ecp) genes; Ecp1, Ecp2, Ecp4 and Ecp5).
M. oryzae Pi-ta / AVR-Pi-ta Fig. 2. Genotype-specific HR in rice seedlings induced by M.grisea carrying AVR-Pita. Sparse HR flecking is seen in Pi-ta-containing rice seedlings (A) Yashiro-mochi and (B) YT14, as expected. In contrast, typical symptoms of rice blast disease are seen in susceptible rice seedlings (C) Nipponbare and (D) YT16. Representative leaves are shown from rice seedlings germinated in plant nutrient medium and infected with avirulent M.grisea strain 4360-R-62 (see Materials and methods for details). Shown at 4 days after inoculation Jia et al. 2000 Pi-ta pi-ta YM YT14 Ni YT16 resistant susceptible Pi-ta dependent resistance response Genotype specific response Intracellular (metalloprotease) direct interaction with Pi-ta Fig. 3. AVR-Pita176 is an elicitor. (A) AVR-Pita polypeptides tested in the transient assay. The white region indicates the putative secretory signal sequence, the gray region indicates the putative pro-protein domain and the hatched region indicates the putative protease motif. The black region indicates the putative mature protein. The number of amino acids missing from the N-terminus is indicated. GUS activity is indicated by ‘+’, whereas decreased GUS activity is indicated by ‘–’. (B) Representative rice seedlings showing GUS activity. Two-leaf Pi-ta (Yashiro-mochi and YT14) and pi-ta (Nipponbare and YT16) seedlings were co-bombarded with 35S/Adh1-6::AVR-Pita176 and 35S::uidA. Leaves were assayed histochemically for GUS activity and cleared in 70% ethanol to visualize GUS staining. (C) RNA gel blot analysis of AVR-Pita expression in the transient assay. YT14 (Pi-ta) and YT16 (pi-ta) were co-bombarded with the 35S/Adh1-6::AVR-Pita176 and 35S::uidA plasmids. Leaf tissue was harvested 2 days after bombardment. Poly(A)+ mRNA was then extracted, blotted to Hybond-N and hybridized with a radiolabeled AVR-Pita176 probe. The AVR-Pita transcript is indicated. Similar loading was verified before blotting by visualizing mRNA in the gel stained with ethidium bromide. Jia et al. 2000
Magnaporthe oryzae is also a root pathogen hydrophobic surface roots a−c, GFP-tagged M. grisea (Guy11) forms classical appressoria (AP) on a hydrophobic surface (a) and simple hyphopodia (HY) and infection pegs (IP) on rice roots (cultivar CO39) (b, c). d−k, Guy11-infected barley (d, e, j, k) and rice (f−i) roots stained with chlorazole black E showing: dark runner hyphae (RH) and simple hyphopodia (d, e); bulbous infection hyphae invading epidermal cells (f); microsclerotia (previously reported in culture). Scale bar, 25 µm. Sesma and Osbourn Sesma and Osbourn
Pigment and cAMP are not required albino cAMP- albino a, Roots of barley seedlings (cultivar Golden Promise) that have been mock-inoculated or infected with the M. grisea wild type (WT) or mutant (mel, cpkA) strains. b, Formation of hyphopodia-like structures (HY) and invasive growth within epidermal cells during the early stages of infection. Scale bars, 25 µm (b), 40 µm (c). cAMP- Sesma and Osbourn
M. oryzae can move systemically from roots to leaves Roots of barley seedlings (cultivar Golden Promise) that have been mock-inoculated or infected with the M. grisea wild type (WT) strain.. c, M. grisea penetrates the stele. Confocal imaging of radial and longitudinal sections of a three-week-old rice seedling (cultivar Nipponbare) infected with GFP-tagged M. grisea (strain Guy11). Scale bars, 25 µm (b), 40 µm (c). Sesma and Osbourn
M. oryzae can move systemically from roots to leaves Pi-CO39(t)-mediated specific disease resistance operates in rice roots a−c, Four-week-old root-infected rice seedlings (cultivar Nipponbare) showing disease symptoms on the leaf (upper box) and collar (lower box) (a). Disease symptoms on the collar (b) and stem (c) with confocal images showing GFP-expressing M. grisea Guy11 in the diseased areas and also in the vascular tissue of the leaf and stem. d−f, Pi-CO39(t)-mediated specific disease resistance operates in rice roots. Confocal microscopy of compatible (d, e) and incompatible (f) interactions. Cultivar, cv. Scale bar, 40 µm. Sesma and Osbourn