Download

1 / 38
460 likes | 1.06k Views
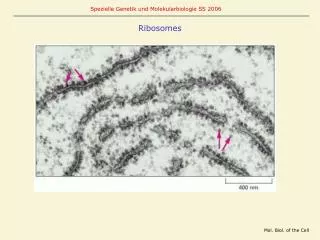
第八章 核糖体 (ribosome). 概述 核糖体的形态结构与存在类型 核糖体的理化性质 核糖体的自组装 核糖体的功能. 核糖体概 述 (形态、大小、数目及分布) 40 年代初,发现细胞内有一种颗粒状结构与蛋白质合成有关。 1955 年, Palade 研究了这种颗粒的性质。曾称之为“ Palade 颗粒”之称。 1958 年, Roberts 根据这种颗粒组成成分的性质,提出了核糖核蛋白体的概念, 简称核糖体或核蛋白体( ribosome ) ,是细胞内蛋白质合成的“工作台”。. 核糖体的功能:合成蛋白质。

E N D
第八章 核糖体(ribosome) 概述 核糖体的形态结构与存在类型 核糖体的理化性质 核糖体的自组装 核糖体的功能
核糖体概 述(形态、大小、数目及分布) 40年代初,发现细胞内有一种颗粒状结构与蛋白质合成有关。 1955年,Palade研究了这种颗粒的性质。曾称之为“Palade颗粒”之称。 1958年,Roberts根据这种颗粒组成成分的性质,提出了核糖核蛋白体的概念,简称核糖体或核蛋白体(ribosome),是细胞内蛋白质合成的“工作台”。
核糖体的功能:合成蛋白质。 • 是细胞最基本的不可缺少的结构:支原体、线粒体、叶绿体中都有。病毒和哺乳动物成熟的红细胞中没有。 • 由rRNA+r蛋白构成。 • 分类:附着核糖体和游离核糖体。 • 分布在细胞内蛋白质合成旺盛的区域。
第一节 核糖体的形态结构与存在类型 核糖体的形态结构 核糖体的存在类型
一、核糖体的形态结构 核糖体上与蛋白质合成有关的结合位点与催化位点 mRNA的结合位点,位于小亚单位。(16SrRNA的3`端有一段顺序同多数原核生物的mRNA(AUG上游3-9个碱基)的核糖体结合位点有互补关系) A位点。与新掺入的氨酰-tRNA的结合位点,又称氨酰基位点,大亚单位和小亚单位都参与。 P位点。与延伸中的肽酰-tRNA的结合位点,又称肽酰基位点,大亚单位和小亚单位都参与。 E位点(exit site)。肽酰转移后与即将释放的tRNA的结合位点,位于大亚单位。
GTP酶的结合位点。催化肽酰tRNA从A位点转移到P位点有关的转移酶,即延伸因子EF-G的结合位点,位于大亚基上。GTP酶的结合位点。催化肽酰tRNA从A位点转移到P位点有关的转移酶,即延伸因子EF-G的结合位点,位于大亚基上。 肽酰转移酶的催化位点。催化氨基酸之间形成肽键的酶(催化P位上肽酰tRNA的羟基与处在A位上氨酰基tRNA的氨基之间形成肽链)。位于大亚基上 。 与蛋白质合成有关的其它起始因子、延伸因子和释放因子的结合位点
二、核糖体的存在类型 • 核糖体单体:无蛋白质合成功能 • 多聚核糖体:合成蛋白质的功能单位 • 游离核糖体:以游离的形式存在于细胞质中 • 附着核糖体:附着于内质网膜表面
多聚核糖体 概念 核糖体在细胞内并不是单个独立地执行功能,而是由多个甚至几十个核糖体串连在一条mRNA分子上高效地进行肽链的合成,这种具有特殊功能与形态结构的核糖体与mRNA的聚合体称为多聚核糖体。 多聚核糖体的生物学意义 细胞内各种多肽的合成,不论其分子量的大小或是mRNA的长短如何,单位时间内所合成的 多肽分子数目都大体相等。 以多聚核糖体的形式进行多肽合成,对mRNA的利用及对其浓度的调控更为经济和有效。
第二节 核糖体的理化性质 主要成分 r蛋白质:35%,核糖体表面 rRNA: 65%,,核糖体内部 基本类型 70S的核糖体:存在于原核细胞、真核细胞的线粒体和叶绿体中。50S+30S。 50S=23SrRNA+5SrRNA+31种r蛋白 30S=16SrRNA+21种r蛋白 80S的核糖体:存在于真核细胞。60S+40S。 60S=28SrRNA+5.8SrRNA+5SrRNA+49种r蛋白 40S=18SrRNA+33种r蛋白 大小亚单位常游离于细胞质基质中,只有当小亚单位与mRNA结合后大亚单位才与小亚单位结合成完整的核糖体。
第三节 核糖体的自组装 蛋白质结合到rRNA上具有先后层次性。 核糖体的重组装是自我装配过程 同一生物中不同种类的r蛋白的一级结构均不相同,在免疫学上几乎没有同源性。 不同生物同一种类r蛋白之间具有很高的同源性,并在进化上非常保守。
16SrRNA的一级结构是非常保守的 16SrRNA的二级结构具有更高的保守性:臂环结构(stem-loop structure) 位于核糖体的A位点和P位点的tRNA都是与16SrRNA配对结合,这些结合部位都处在16SrRNA高度保守序列上。
第四节 核糖体的功能 • 核糖体在蛋白质生物合成中执行两项重要功能: 1、使mRNA不断与tRNA分子结合 2、控制生长中的肽链
真核细胞中蛋白质合成的不同点 • 核糖体小亚单位与mRNA的5‘端的cap识别并结合在一起,然后沿mRNA移动至遇到起始密码AUG,此过程称为扫描。 • 参与翻译的起始因子是eIF系列。 • 形成起始复合物的氨基酸是蛋氨酸而不是甲酰蛋氨酸。 • 与原核细胞延长因子EF-Tu和EF-G对应的真核细胞延长因子为EF1和EF2。
蛋白质合成过程中很多重要步骤与50S核糖体大亚单位相关蛋白质合成过程中很多重要步骤与50S核糖体大亚单位相关 1、依赖延伸因子Tu的氨酰tRNA的结合; 2、延伸因子G介导的转位作用; 3、依赖于起始因子2的fMet-tRNA的结合; 4、依赖于释放因子的蛋白合成终止作用; 5、应急因子(营养缺乏条件下促代谢水平迅速下降的因子)与核糖体结合产生ppGpp(p)阻断蛋白合成等。
核糖体蛋白质与rRNA的功能分析 核糖体上具有一系列与蛋白质 合成有关的结合位点与催化位点 在蛋白质合成中肽酰转移酶的活性研究
在蛋白质合成中肽酰转移酶的活性研究 核糖体蛋白 在核糖体中rRNA是起主要作用的结构成分 r蛋白质的主要功能
核糖体蛋白 很难确定哪一种蛋白具有催化功能:E.coli中核糖体蛋白突变甚至缺失对蛋白质合成并没有表现出“全”或“无”的影响。 多数抗蛋白质合成抑制剂的突变株,并非由于r蛋白的基因突变而往往是 rRNA基因突变。 在整个进化过程中rRNA的结构比核糖体蛋白的结构具有更高的保守性。
在核糖体中rRNA是起主要作用的结构成分 具有肽酰转移酶的活性; 为tRNA提供结合位点(A位点、P位点和E位点); 为多种蛋白质合成因子提供结合位点; 在蛋白质合成起始时参与同mRNA选择性地结 合以及在肽链的延伸中与mRNA结合; 核糖体大小亚单位的结合、校正阅读(proofreading)、无意义链或框架漂移的校正、以及抗菌素的作用等都与rRNA有关。
r蛋白质的主要功能 对rRNA 折叠成有功能的三维结构是十分重要的; 在蛋白质合成中, 某些r蛋白可能对核糖体的构象 起“微调”作用; 在核糖体的结合位点上甚至可能在催化作用中, 核 糖体蛋白与rRNA共同行使功能。
RNA在生命起源中的地位及其演化过程 RNA的功能 DNA代替了RNA的遗传信息功能 蛋白质取代了绝大部分RNA酶的功能
RNA的功能 三种生物大分子,只有RNA既具有信息载体 功能又具有酶的催化功能。因此,推测RNA 可能是生命起源中最早的生物大分子。 核酶(ribosome):具有催化作用的RNA。 由RNA催化产生了蛋白质
核糖很容易由甲烷合成,脱氧核糖需由核糖还原而来,因此生命的最早形式可能是膜围绕的一套具有自我复制能力的分子体系和简单的物质及能量供应体系,遗传物质载体是RNA。核糖很容易由甲烷合成,脱氧核糖需由核糖还原而来,因此生命的最早形式可能是膜围绕的一套具有自我复制能力的分子体系和简单的物质及能量供应体系,遗传物质载体是RNA。 • 后来出现了RNA催化形成的蛋白质来取代大部分RNA酶的部分功能。 • 再后来DNA代替了RNA的遗传信息功能。
DNA代替了RNA的遗传信息功能 DNA双链比RNA单链稳定; DNA链中胸腺嘧啶代替了RNA链中的尿嘧啶,使之易于修复。有利于信息的稳定遗传。
蛋白质取代了绝大部分RNA酶的功能 蛋白质化学结构的多样性与构象的多变性; 与RNA相比,蛋白质能更为有效地催化多种生化反应,并提供更为复杂的细胞结构成分,逐渐演化成今天的细胞。
第五节 异常情况下核糖体的变化 • 多聚核糖体的解聚:指多聚核糖体分散为单体,孤立得分散在细胞质中或附着在糙面内质网膜上。 多聚核糖体的解聚伴随着内源性蛋白质生成的减少。 幼红细胞和网织红细胞:负责合成血红蛋白,细胞质中充满多聚核糖体。 当成熟为红细胞:核糖体消失。 分化低的肿瘤细胞和胚胎细胞:细胞质中充满多聚核糖体。 细胞有丝分裂阶段:蛋白质合成明显下降,多聚核糖体解聚。
脱粒:附着在糙面内质网上的核糖体脱落下来,散在细胞质中,核糖体往往同时解聚为单体。脱粒:附着在糙面内质网上的核糖体脱落下来,散在细胞质中,核糖体往往同时解聚为单体。 核糖体脱粒并解聚意味着输出蛋白质形成减少。 维生素C缺乏引起的坏血病中,成纤维细胞中糙面内质网膜上的多聚核糖体解聚为单体,但不脱落,蛋白质合成减少。 创伤愈合的成纤维细胞,糙面内质网上附着许多核糖体,蛋白质合成增强。 CCl4中毒引起的肝损伤,出现脱粒,蛋白质合成急剧下降
第六节 影响蛋白质合成的药物 一、血红素对血红蛋白合成的调节 蛋白激酶 + eIF-2磷酸化, 失活 cAMP 起始因子2激酶 - 高铁血红素 保护eIF-2不失活,有 利于血红蛋白的合成
eIF-2磷酸 化失活 抑制蛋白 质合成 eIF-2激酶 诱导合成 二、干扰素对蛋白质合成的调节 干扰素 2´,5´-寡聚腺 苷酸合成酶 2´,5´-寡 聚腺苷酸 + 内切核酸酶 ATP 降解mRNA,减 少蛋白质合成
三、抗生素对蛋白质生物合成的影响 1、四环素类药物:抑制氨酰-tRNA与原核细胞核糖体的结合,抑制多种细菌的蛋白质合成; 2、氯霉素:与原核细胞核糖体的50S亚基结合,阻断肽键形成;高浓度时对哺乳动物线粒体内核糖体50S亚基也有作用; 3、链霉素与卡那霉素:与原核细胞核糖体30S亚基结合,改变核糖体构象,引起读码错误,合成错误蛋白质; 4、嘌呤霉素:结构与酪氨酰-tRNA末端相似,带有游离氨基,可取代氨酰-tRNA进入核糖体受位,使正在延长的肽链转移到嘌呤霉素的氨基上,异常肽链从核糖体释放,阻止肽链延长。对真核细胞和原核细胞都有作用。
5、放线菌酮:抗真菌,抑制真核细胞生物核糖体上的多肽转移酶,作用于60S核糖体抑制蛋白质合成的起始和延长。5、放线菌酮:抗真菌,抑制真核细胞生物核糖体上的多肽转移酶,作用于60S核糖体抑制蛋白质合成的起始和延长。 6、大环内酯类抗生素:红霉素、克拉霉素、麦迪霉素、阿奇霉素、乙酰螺旋霉素、交沙霉素等。 通过阻断转肽作用和mRNA的移位,抑制蛋白质的合成。 耐药机制:抗生素与核糖体结合部位改变。耐药菌合成甲基化酶,使50S亚基的23SrRNA腺嘌呤甲基化,导致抗生素不能与核糖体结合部位结合。
本 章 重 点 • 核糖体的结构成分。核糖体的功能。 • 蛋白质的合成过程。 • 核糖体上有哪些活性位点?在多肽合成中起什么作用? • 何谓多聚核糖体?以多聚核糖体的形式行使生物学功能的意义是什么? • RNA是最早出现的简单生命体中的生物大分子。