Download

1 / 56
560 likes | 685 Views
Within-individual variation in bullfrog vocalizations Implications for a vocally mediated social recognition system. Mark A. Bee J. Acoust. Soc. Am. 116 (6). (Fischer et al. , 2002 ). (Robertson, 1996 ). (Weary et al. , 1990). (Polakow, 1997 ). Introduction.

E N D
Within-individual variation in bullfrog vocalizations Implications for a vocally mediated social recognition system Mark A. Bee J. Acoust. Soc. Am. 116 (6)
(Fischer et al., 2002 ) (Robertson, 1996 ) (Weary et al., 1990) (Polakow, 1997 ) Introduction • Vocal communication plays important roles in social recognition • Allowing animals to direct appropriate behaviors toward other individuals (Bee and Gerhardt, 2001a) (Myrberg et al., 1993)
Introduction • animals behaviorally discriminate based on the learned recognition of individually distinct acoustic signals in different behavioral contexts, such as • kin recognition (Beecher, 1991; Balcombe, 1990; Aubin and Jouventin, 2002) • mate recognition (Robertson, 1996; Lengagne et al., 2000) • territorial neighbor recognition (Brooks and Falls, 1975a; Myrberg and Riggio, 1985; Bee and Gerhardt, 2001b, c, 2002) • individual recognition (Beecher et al., 1996; Cheney and Seyfarth, 1999; Gentner and Hulse, 1998, 2000)
Introduction • However, relatively few studies have sought to understand the mechanisms and evolution of vocally mediated social recognition by systematically investigating the acoustic and perceptual bases of recognition either within or across taxa (Beecher, 1991; Gentner and Hulse, 1998, 2000; Bee and Gerhardt, 2001a, b, 2002; Bee, 2001, 2003 )
Introduction • Clearly an important aspect of acoustically mediated recognition systems involves the patterns of variation in acoustic signals that identify individuals • Only after characterizing the potential for specific acoustic properties to identify individuals can researchers investigate in an informed way the perceptual basis of recognition using playback experiments.
Introduction • A study of a vocally mediated social recognition system in a frog • Bullfrog lower levels of aggression in response to neighbor’s advertisement calls from the neighbor’s territory (Davis, 1987)
Introduction • Bee and Gerhardt (2001a) investigated the acoustic basis of neighbor-stranger discrimination in bullfrogs • The fundamental frequency exhibited the greatest among-individual variation relative to within-individual variation
Introduction • Using the habituation-discrimination paradigm would allow males to learn to recognize acoustic properties • changes in the fundamental frequency elicited a recovery of aggressive responses • territory residents use features related to call pitch to identify neighbors as familiar. (Bee and Gerhardt ~2001b, c, 2002; Bee, 2003)
Introduction • Based on the acoustic analyses and field playback experiments, the ‘‘just-meaningful difference’’ (JMD) (Nelson and Marler, 1990; Gerhardt, 1992) in fundamental frequency was between 5-10% • this conclusion was provisional for two important reasons (Bee and Gerhardt, 2001a, b, c)
Introduction • First, variation in several acoustic properties were reported, but not describe • including fundamental frequency, that occurred among the separate notes within the multiple note advertisement calls • This note-to-note variation could increased overlap of the signal properties of different individuals’ calls. • Second, the synthetic stimuli were invariant • the note-to-note within-individual variation present in multiple-note advertisement calls was not incorporated in the perceptual learning and discrimination task. (Bee and Gerhardt, 2001a)
Introduction • Thus, an important and unanswered question - within-individual variation in multiple-note advertisement calls for the behavioral discrimination thresholds
Introduction • The main objective of this study was to determine the variation in multiple-note calls can either constrain or provide additional cues for neighbor recognition
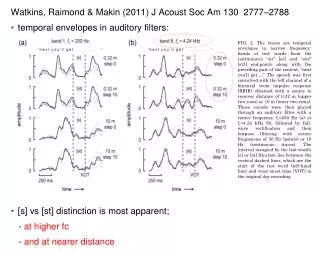
Method- study organism • During their breeding season, male bullfrogs • establish and defend territories • most advertisement calls consist of four to seven notes • The individual notes are about 700 ms in duration, are separated by intervals of about 500 ms • periodicity varies inversely with body size and corresponds to the fundamental frequency of the call (Howard, 1978) (Capranica, 1965; Bee and Gerhardt, 2001a)
Method- study organism • The frequency spectrum of a call note consists of a series of harmonics. • The lower frequency peak is between 200 and 400 Hz; a broader, higher frequency band occurs between 1000 and 2000 Hz
HHb PDR-1000 DAT recorder Method- recording of vocalization • Between May and August 1998, 25 territorial male bullfrogs • between 2300 and 0400 hours, at least five multiple-note advertisement calls • sampling rate532 kHz, 1–2 m from a subject • the air and water temperatures at the frog’s position • Each subject had its SVL, measured on a night prior to that of recording (following Bee and Gerhardt 2001a)
Method- Acoustical analysis • spectral properties that were measured • dominant frequency (DF) • secondary frequency (SF) • the amplitude of the dominant frequency relative to the secondary frequency (RA) • the ratio of secondary frequency to dominant frequency (frequency ratio)
Method- Acoustical analysis • Three gross-temporal properties were measured from oscillograms • note duration (ND) • rise time (RT) • fall time (FT)
Method- Acoustical analysis • Using an oscillogram with an expanded time base, fundamental frequency was determined
Method- statistical analysis • ANOVA • MANOVA • Discriminant function analysis (DFA) + PCA(Nelson and Marler, 1990; Weary et al., 1990; Hauser, 1991; Insley, 1992; Bee et al., 2001; Fischer et al., 2002) • Discrimination threshold, JMDs (Nelson and Marler, 1990; Gerhardt, 1992)
Result- Sources and patterns of variation • R2 values that ranged from 0.59 to 0.95. • Variation among individuals was significantly greater than that within individuals
note interaction individual
posthoc comparisons revealed that • fundamental frequency and dominant frequency were significantly lower in the first note • secondary frequency was significantly lower at the beginning and end, and notes at the beginning of a call were significantly lower than notes at the end • the amplitude of the dominant frequency relative to that of the secondary frequency was significantly lower in the first note • the dominant frequency and secondary frequency are harmonically related to the fundamental
there was a slight trend for frequency ratio to be higher in notes from the middle of a call. • Together, the results for secondary frequency and frequency ratio suggest that the spectral energy in the upper mode of the frequency spectrum shifted to slightly higher harmonics in the middle of the calls
Average note duration was constant within a call • there were significant differences in rise times and fall times • Posthoc comparisons indicated that rise times increased, and fall times decreased
spectrum properties • tended to vary little: FF(3%), DF(3%), SF(7%), and frequency ratio (4%) • The exception was relative amplitude(18%) • temporal properties • note duration was constant across multiple notes, varying less than 1% • more variation was observed for rise time(25%) and fall time (24%)
note interaction individual
most individuals had secondary frequencies that were either relatively constant or increased slightly • however, at least a few individuals had average secondary frequencies that were considerably more variable • it is unlikely due to changes in the positions of some individuals relative to the microphone • Hence, the patterns of changes in the frequency of peak sound energy in the upper mode of the bimodal frequency spectrum that occurred among notes within multiple note calls differed reliably
The reliable individual differences in note-to-note variation were even more pronounced for note duration • less than 0.5% of the variation was explained by note position • the duration of notes was nearly constant within some individuals while for other individuals note duration changed by 100–200 ms
Result- relative variation among acoustic properties • There were significant differences in the variability among call properties • within-individual CV (Wilks’ λ<0.01, F7,18=423.1, P <0.0001 • within-individual ranges (Wilks’ λ<0.01, F7,18=223.7, P<0.0001 )
The DFA successfully assigned 283 of the 375 recorded call notes (75.5%) to the correct individual • which is significantly higher than the classification success of 4% expected by randomly assigning notes to individuals (Fisher’s exact test, P ,0.01). • Therefore, individuals can be statistically distinguished by their multiple-note advertisement calls.
resident familiar stranger familiar stranger reject alarm • there would be a selective advantage to having a behavioral discrimination threshold that leads to high correct rejection rates and low false alarm rates. fail fail correct correct
Discussion- Implications of within-individual variation • Bee and Gerhardt (2001a) reported several acoustic properties of bullfrog , the results reported above extend this previous work in several important ways. • First, the results describe the systematic patterns of variation in call properties present among notes within an individual’s calls. • Second, the results from the present study suggest that there may be significant among-individual differences in the patterns of within individual variation
Third, the results show that, with the exception of relative amplitude, fine-temporal and spectral call properties exhibited significantly lower magnitudes of variation within individuals • Finally, fundamental frequency and dominant frequency exhibited the most reliable among-individual differences
Discussion- Individual discrimination based on ‘‘pitch’’ • As in some songbirds, male bullfrogs appear to use acoustic properties to discriminate among familiar and unfamiliar vocalizations. (Brooks and Falls, 1975b; Nelson, 1989) • Bee and Gerhardt (2001b,c) demonstrated that a 5–10% change in fundamental frequency was sufficient to elicit significant levels of recovery of the habituated aggressive response. • Hence, the JMDs for fundamental frequency and correlated spectral properties of 4%–6%
Psychophysical estimates of frequency discrimination are available for only one frog, the South African clawed frog, Xenopus leavis (Elepfandt et al., 2000) • Frequency difference limens for pure tones measured were approximately 5% in the hearing range of the amphibian papilla, and 2.4%–6% in the hearing range of the basilar papilla.
The current estimates of JMDs for fine-temporal or spectral properties of bullfrog advertisement calls may, therefore, approach the absolute limits of the anuran auditory system.
Discussion- Individual discrimination based on other acoustic properties • At present, there have been only limited efforts to investigate the use of other call properties as recognition cues in bullfrogs. • Other properties of advertisement calls also have some potential to statistically distinguish and therefore we might expect behavioral discrimination to be based on multiple dimensions of signal variability. • Although the sample sizes were small, found no evidence in their habituation-discrimination experiments that 10% decreases in note duration and internote interval or 10% increases in rise and fall times elicit recovery of habituated aggressive responses. (Bee and Gerhardt, 2001a; Simmons, 2004), (Bee and Gerhardt 2001)
Novel stimuli having differences in the harmonic fine-structure ~presence/absence and relative amplitudes of harmonic components!, but a constant fundamental frequency, also failed to elicit recovery of habituated aggressive responses ~Bee and Gerhardt, 2001c!. • On the one hand, these previous findings are consistent with the relatively higher estimated JMDs for rise time, fall time, and relative amplitude ~Table IV! in comparison to those for fine-temporal and spectral call properties. • On the other hand, however, if note duration is used as a recognition cue, then these results suggest that the JMD for note duration used by receivers is higher than that estimated here. • Estimating JMDs for note duration is complicated by the relatively large among-individual differences in the patterns of note-to-note variation. • Clearly, more work is needed to determine whether receivers use gross temporal call properties as recognition cues.
Three properties of bullfrog advertisement calls, for which the patterns of among-individual and within-individual variation have not been investigated thoroughly, are internote interval, note duty cycle, and phase spectrum. • At a constant note duty cycle, a 10% decrease in internote interval ~and note duration! failed to elicit response recovery ~Bee and Gerhardt, 2001c!. • When note duration was held constant and the duty cycle was increased 20% ~from 0.5 to 0.7! by decreasing internote interval by 57% ~700 to 300 ms!, half of the subjects responded aggressively to the novel stimulus @only 25% responded to the control stimulus ~Bee and Gerhardt, 2001c!#.
None of the subjects responded to a novel stimulus having a 10% increase in duty cycle ~from 0.5 to 0.6! effected by a 33% decrease in internote interval ~700 to 467 ms!. • In a field playback test examining habituation and recovery of evoked advertisement calling, Hainfeld et al. ~1996! found some evidence to suggest that territorial male bullfrogs might behaviorally discriminate among advertisement calls differing in phase spectrum based on differences in waveform periodicity. • If there were reliable individual differences in the phase spectra of advertisement calls that were reliably transmitted through the environment across distances that separate individuals, then the phase spectrum of a neighbor’s calls might also function as an additional acoustic cue for recognition. • This seems unlikely, however, as Hainfeld et al. ~1996! argued that phase spectrum was not a particularly salient cue for discriminating among advertisement calls in the natural environment.
Conclusions • Surprisingly little effort has been made to integrate, either within or across species, an understanding of the underlying acoustic and perceptual bases of vocally mediated social recognition. • The present study represents the latest effort to better understand both the acoustic and perceptual bases of vocally mediated neighbor recognition in North American bullfrogs (Bee and Gerhardt, 2001a,b,c, 2002, Bee, 2001, 2003)
The ultimate goal of acoustical and statistical analyses like those reported here should be to relate patterns of signal variation both to the JMD required to elicit evolutionarily adaptive behavioral responses from receivers, as well as to the ‘‘just-noticeable difference’’ ~JND! of the receiver’s auditory system. • The habituation-discrimination paradigm represents a powerful experimental tool for providing behavioral estimates of receivers’ JMDs under natural conditions.
Psychophysical estimates of the JMDs of acoustic signal properties are generally still lacking for frogs. • Future psychophysical experiments with bullfrogs could provide much needed insights into the mechanisms of vocal recognition in bullfrogs (Elepfandt et al., 2000)
Discussion- relevance of estimating JMDs from patterns of individual variation in signals • Acoustical and statistical analyses of signals cannot determine a receiver’s discrimination thresholds, which instead requires experimentation. • Therefore, it is important for readers to keep in mind that estimating JMDs from the degree of overlap in the distributions of among-individual and within-individual differences in multiple note calls represents one way to generate hypotheses about a receiver’s behavioral discrimination threshold that can be tested in future studies. • The value of deriving estimated JMDs in this way is that it provides a reasonable starting point from which to assess actual JMDs in field playback experiments.