Download
1 / 1
10 likes | 90 Views
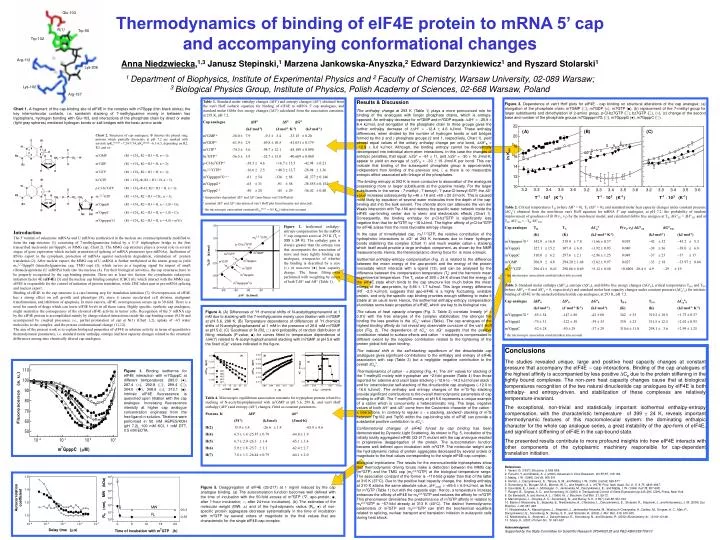
Glu-103. N(1). Trp-56. Trp-102. Arg-112. Lys-206. Lys-162. (A). (B). Arg-157. (C).

E N D
Glu-103 N(1) Trp-56 Trp-102 Arg-112 Lys-206 Lys-162 (A) (B) Arg-157 (C) Figure 3. Dependence of van’t Hoff plots for eIF4E - cap binding on structural alterations of the cap analogue: (a) elongation of the phosphate chain, m7GMP (), m7GDP (□), m7GTP (■), (b) replacement of the 7-methyl group for larger substituents and dimethylation of 2-amino group, p-Cl-bz7GTP (), bz7GTP (), (○), (c) change of the second base and number of the phosphate groups m7Gppppm7G (), m7GpppG (●), m7GpppC (). Chart 2. Structures of cap analogues; denotes the phenyl ring; protons which partially dissociate at pH 7.2 are marked with asterisk (pKaN(1)-H ~7.24-7.54, pKaphosph ~6.1-6.5, depending on R2, R3, and n): m7GMP (R1 = CH3, R2 = R3 = H, n = 1); m7GDP (R1 = CH3, R2 = R3 = H, n = 2); m7GTP (R1 = CH3, R2 = R3 = H, n = 3); bz7GTP (R1 = CH2-, R2 = R3 = H, n = 3); p-Cl-bz7GTP (R1 = CH2--Cl, R2 = R3 = H, n = 3); m32,2,7GTP (R1 = CH3, R2 = R3 = CH3, n = 3); m7GpppG (R1 = CH3, R2 = R3 = H, n = 3, B = G); m7GpppC (R1 = CH3, R2 = R3 = H, n = 3, B = C); m7Gppppm7G (R1 = CH3, R2 = R3 = H, n = 4, B = m7G) Table 2. Critical temperatures TH (where H = 0), TS (S = 0), and standard molar heat capacity changes under constant pressure (Cp) obtained from the non-linear van't Hoff equation for mRNA 5' cap analogues, at pH 7.2, the probability of random improvement of goodness of fit P(1, 2) by the non-linear model, and calculated Gibbs free energies at TS, GTS = HTS, and at TH, GTH = THSTH. Cap analogue TH TSCp P(1, 2) GTSGTH (K) (K) (kJ·mol-1·K-1) (kJ·mol-1) (kJ·mol-1) m7Gppppm7Ga 342.0 16.0 318.0 7.8 +1.66 0.57 0.019 42 32 43.2 5.3 m7GpppG 327.1 15.2 307.4 6.0 +1.92 0.93 0.040 38 36 39.0 6.8 m7GpppC 310.0 6.2 297.6 2.1 +2.96 1.25 0.049 37 25 37 17 m7GMP306.9 4.8 294.20 1.68 +2.62 0.97 0.027 33 18 33.97 0.84 m32,2,7GTP 296.41 0.43 290.86 0.69 +5.12 0.48 <0.0001 28.4 4.9 29 19 a the microscopic association constant taken into account Figure 2. Isothermal enthalpy-entropy compensation for the mRNA 5’ cap congener series at 293 K (Tc = 399 24 K). The enthalpy gain is always greater than the entropy loss that accompanies the association of more and more tightly binding cap analogues, irrespective of whether the binding is described by a zero (○) or non-zero (●) heat capacity change. The linear fitting was performed with weighting by errors of both TS and H (Table 1). Table 3. Standard molarenthalpy (H0), entropy (S0), and Gibbs free energy changes (G0), critical temperatures TH 0 and TS 0 (where H0 = 0 and S0 = 0, respectively) and standard molar heat capacity changes under constant pressure (Cp0) for intrinsic binding of eIF4E to the unstacked dinucleotide cap analogues, at 293 K, pH 7.2. Cap analogue H0S0G0 TH 0 TS 0Cp0 (kJ·mol-1) (J·mol-1·K-1) (kJ·mol-1) (K) (K) (kJ·mol-1·K-1) m7Gppppm7Ga -85 54 -147 88 -42 60 342 35 319.2 18.5 +1.73 0.57 m7GpppG -75 31 -122 58 -39 35 330 23 311.5 12.5 +2.01 0.93 m7GpppC -52 28 -50 29 -37 29 310.6 11.8 298.1 3.6 +2.99 1.25 a the microscopic association constant taken into account Figure 4. (A) Differences of 1H chemical shifts of N-acetyltryptophanamid at 1 mM due to stacking with the 7-methylguanine moiety upon titration with m7GMP at pH 5.6, 298 K; (B) Temperature dependence of differences of 1H chemical shifts of N-acetyltryptophanamid at 1 mM in the presence of 29.8 mM m7GMP at pH 5.6; (C) Goodness of fit (R2, ) and probability of random distribution of fitting residuals (P-value, ■) for curves fitted to temperature dependence of H(7) related to N-acetyl-tryptophanamid stacking with m7GMP at pH 5.6 with the fixed Cp values indicated in the figure. Figure 1. Binding isotherms for eIF4E interaction with m7GpppC at different temperatures: 280.0 (●), 287.4 (○), 292.8 (), 299.4 (), 304.0 (), 308.6 (□), 313.1 (■). Intrinsic eIF4E fluorescence is quenched upon titration with the cap analogue. Increasing fluorescence intensity at higher cap analogue concentration originates from the free ligand in solution. Titrations were performed in 50 mM HEPES/KOH (pH 7.2), 100 mM KCl, 1 mM DTT, 0.5 mM EDTA. Table 4. Microscopic equilibrium association constants for tryptophan protons related to stacking of N-acetyltryptophanamid with m7GMP at pH 5.6, 298 K, and van't Hoff enthalpy (H) and entropy (S) changes, fitted as constant parameters. Protons K HS (M-1) (kJ/mol) (J/molK) H(2) 15.9 3.8 -26.6 1.8 -65.8 4.6 H(4) 6.5 1.6 -25.87 0.78 -64.0 1.9 H(5) 6.7 2.0 -26.3 1.4 -65.1 3.4 H(6) 5.9 1.8-25.7 1.1 -62.4 2.7 H(7) 7.8 1.3 -26.44 0.79 -64.1 2.0 Figure 5. Deaggregation of eIF4E (33-217) at 1 mg/ml induced by the cap analogue binding. (a) The autocorrelation function becomes well defined with the time of incubation with the 50-fold excess of m7GTP (▽, apo-protein; ■, after 1-hour incubation; ○, after 24-hour incubation). (b) The estimates of the molecular weight (MW, △) and of the hydrodynamic radius (Rh, ●) of non-specific protein aggregates decrease systematically in the time of incubation with m7GTP by several orders of magnitude to the final values that are characteristic for the single eIF4E-cap complex. Thermodynamics of binding of eIF4E protein to mRNA 5’ cap and accompanying conformational changes Anna Niedzwiecka,1,3 Janusz Stepinski,1 Marzena Jankowska-Anyszka,2 Edward Darzynkiewicz1 and Ryszard Stolarski1 1 Department of Biophysics, Institute of Experimental Physics and2Faculty of Chemistry, Warsaw University, 02-089 Warsaw; 3Biological Physics Group, Institute of Physics, Polish Academy of Sciences, 02-668 Warsaw, Poland Table 1. Standard molarenthalpy changes (H) and entropy changes (S) obtained from the van't Hoff isobaric equation for binding of eIF4E to mRNA 5' cap analogues, and standard molar Gibbs free energy changes (G) calculated from the association constants, at 293 K, pH 7.2. Cap analogue HSG (kJ·mol-1) (J·mol-1·K-1) (kJ·mol-1) m7GMPa-36.0 7.9 -9.3 3.4 -33.15 0.20 m7GDPb -61.9 2.9 -69.8 10.5 -41.031 0.179 m7GTPb -74.3 3.6 -98.7 12.1 -45.109 0.090 bz7GTPb -56.5 3.8 -52.7 13.0 -40.669 0.060 p-Cl-bz7GTPb -38.5 4.6 +16.7 15.5 -42.94 0.21 m32,2,7GTPa -16.6 2.5 +40.3 12.7 -28.94 1.36 m7Gppppm7Ga, c -81 54 -136 88 -41.377 0.146 m7GpppGa -65 31 -91 58 -38.555 0.152 m7GpppCa -50 28 -45 29 -36.42 0.40 a temperature-dependent H and S (non-linear van’t Hoff plot) b constant H and S (deviation of van’t Hoff plot from linearity not detected) c the microscopic association constant (Kasmicro = 0.5 Kas) taken into account Results & Discussion The enthalpy change at 293 K (Table 1) plays a more pronounced role for binding of the analogues with longer phosphate chains, which is entropy-opposed. An enthalpy decrease for m7GMP and m7GDP equals: H = 25.9 8.4 kJ/mol, and elongation of the phosphate chain to three groups gives the further enthalpy decrease of H = 12.4 4.6 kJ/mol. These enthalpy differences, when divided by the number of hydrogen bonds or salt bridges formed by the and phosphate groups (2 and 1, respectively, Chart 1), yield almost equal values of the unitary enthalpy change per one bond, HB12.8 9.6 kJ/mol. Although, the binding entropy cannot be theoretically decomposed into individual atom-atom interactions, in this case the changes of entropic penalties, that equal: S = 61 11, and S = 30 16 J/molK, appear to yield an average of SB30 19 J/molK per bond. This can indicate that binding of the subsequent phosphate group is approximately independent from binding of the previous one, i. e. there is no measurable entropic effect associated with linkage of the phosphates. The binding entropy at 293 K is more conducive to association of the analogues possessing more or larger substituents at the guanine moiety. For the larger substituents in the series : 7-methyl-, 7-benzyl-, 7-para-Cl-benzyl-GTP, the S value increases subsequently by +46 18 and +69 20 J/molK. This is caused most likely by expulsion of several water molecules from the depth of the cap-binding slot into the bulk solvent. The chloride atom can attenuate the van der Waals interaction with Trp-166 and destroy the specific water network inside the eIF4E cap-binding center due to steric and electrostatic effects (Chart 1). Consequently, the binding enthalpy for p-Cl-bz7GTP is significantly less negative than that for bz7GTP, by ~18 kJ/mol. The higher affinity of p-Cl-bz7GTP for eIF4E arises from the more favorable entropy change. In the case of trimethylated cap, m32,2,7GTP, the relative contribution of the hydrophobic interactions to G at 293 K increases due to fewer hydrogen bonds stabilizing the complex (Chart 1) and much weaker cation- stacking which itself would provide a large enthalpic component, as shown by the NMR measurements. Hence, the thermodynamic driving force for is more entropic. Isothermal enthalpy-entropy compensation (Fig. 2) is related to the difference between the mean energy of the apo-protein and the energy of the protein microstate which interacts with a ligand (13), and can be analysed by the difference between the compensation temperature (Tc) and the harmonic mean experimental temperature. The Tc value of 399 24 K shows that the energy of the eIF4E state which binds to the cap structure lies much below the mean energy of the apo-protein, by 9.66 1.7 kJ/mol. This large energy difference (RT ~2.5 kJ/mol), suggests that apo-eIF4E is a highly fluctuating, unstable protein, and only the specific cap binding provides enough stiffening to make it stable at an usual level. Hence, the isothermal enthalpy-entropy compensation elucidates some basic properties of eIF4E, which are key to its biological role. The values of heat capacity changes (Fig. 3, Table 2) correlate linearly (r2 = 0.91) with the free energies of the complex stabilization: the stronger the binding the less positive is the Cp value (Table). The cap analogues of the highest binding affinity do not reveal any observable curvature of the van't Hoff plot (Fig. 2). The dependence of Cp on G suggests that the positive contribution related to surface effects and cation - stacking is compensated to different extent by the negative contribution related to the tightening of the protein global fold upon binding. The induced shift in the self-stacking equilibrium of the dinucleotide cap analogues gives significant contributions to the enthalpy and entropy of eIF4E association with cap (Table 3) but a negligible negative contribution to the overall Cp°. Thermodynamics of cation - stacking (Fig. 4). The H values for stacking of the 7-methylG moiety with tryptophan are ~2-fold greater (Table 4) than those reported for adenine and uracil base stacking (12.6 to 14.2 kJ/mol per stack ) and for intramolecular self-stacking of the dinucleotide cap analogues (12.0 to 16.6 kJ/mol). The enthalpy and entropy changes of the m7G-Trp stacking provide significant contributions to the overall thermodynamic parameters of cap binding to eIF4E. The 7-methylG moiety at pH 5.6 represents a unique example of a cation which is concurrently a heteroaromatic ring. The large, negative values of both H and S come from the Coulombic character of the cation - interactions. In contrary to regular - stacking, sandwich stacking of m7G between Trp102 and Trp56 within the cap-binding site of eIF4E can provide a substantial positive contribution to Cp. Conformational changesof eIF4E forced by cap binding has been demonstrated by Dynamic Light Scattering. As shown in Fig. 5, incubation of the initially totally aggregated eIF4E (33-217) mutant with the cap analogue resulted in progressive deaggregation of the protein. The autocorrelation function became well defined upon incubation with m7GTP. The molecular weight and the hydrodynamic radius of protein aggregates decreased by several orders of magnitude to the final values corresponding to the single eIF4E-cap complex. Biological implications. The results for the mononucleotide triphosphates show that thermodynamic driving forces make a distinction between the MMG cap (m7GTP) and the TMG cap (m32,2,7GTP) at the biological temperature range. The association constant of the former is ~110-fold greater than that of the latter at 310 K (37C). Due to the positive heat capacity change, the binding enthalpy at 310 K attains the same absolute value, H310K = +69.6 6.9 kJ/mol, as that for m7GTP (Table 1) but with the opposite sign. Hence, a temperature increase enhances the affinity of eIF4E for m32,2,7GTP and reduces the affinity for m7GTP. This phenomenon diminishes the predominance of m7GTP affinity in relation to m32,2,7GTP to ~57-fold already at 313 K (40C). The distinct thermodynamic parameters of m7GTP and m32,2,7GTP can shift the biochemical equilibria related to splicing, nuclear transport and translation initiation in eukaryotic cells during heat shock. Chart 1.A fragment of the cap-binding site of eIF4E in the complex with m7Gppp (thin black sticks); the key intermolecular contacts, i.e. sandwich stacking of 7-methylguanine moiety in between two tryptophans, hydrogen bonding with Glu-103, and interactions of the phosphate chain by direct or water (light gray spheres) mediated hydrogen bonds or salt bridges with the basic amino acids. Introduction The 5’ termini of eukaryotic mRNAs and U snRNAs synthesized in the nucleus are cotranscriptionally modified to form the cap structure (1) consisting of 7-methylguanosine linked by a 5’-5’ triphosphate bridge to the first transcribed nucleoside (m7GpppN, or MMG cap, Chart 2). The MMG cap structure plays a pivotal role in several stages of gene expression which include promotion of splicing of mRNA precursors (pre-mRNAs), facilitation of RNAs export to the cytoplasm, protection of mRNAs against nucleolytic degradation, stimulation of protein translation (2). After nuclear export, the MMG cap of U snRNA is further methylated at the amino group to yield m32,2,7GpppN (trimethylguanosine cap, TMG cap) (3), which serves as a targeting signal for import of small ribonucleoproteins (U snRNPs) back into the nucleus (4). For their biological activities, the cap structures have to be properly recognized by the cap-binding proteins. There are at least two factors: the cytoplasmic eukaryotic initiation factor 4E (eIF4E) (5) and the nuclear cap binding complex (CBC) (6), which interact with the MMG cap. eIF4E is responsible for the control of initiation of protein translation, while CBC takes part in pre-mRNA splicing and nuclear export. Binding of eIF4E to the cap structure is a rate-limiting step for translation initiation (7). Overexpression of eIF4E has a strong effect on cell growth and phenotype (8), since it causes accelerated cell division, malignant transformation, and inhibition of apoptosis. In most cancers, eIF4E overexpression occurs up to 30-fold. There is a need for search of drugs which can lower eIF4E levels in all those cases. Highly specific, synthetic cap analogues, might neutralize the consequences of the elevated eIF4E activity in tumor cells. Recognition of the 5’ mRNA cap by the eIF4E protein is accomplished mainly by charge-related interactions inside the cap-binding center (9,10) and accompanied by coupled processes, i.e., partial protonation of cap at N(1) (Chart 1,2), uptake of ~65 water molecules to the complex, and the protein conformational change (11,12). The aim of the present work is to explain biological properties of eIF4E in solution activity in terms of quantitative thermodynamic parameters, i.e. standard molar enthalpy, entropy and heat capacity changes related to the structural differences among nine chemically altered cap analogues. Conclusions The studies revealed unique, large and positive heat capacity changes at constant pressure that accompany the eIF4E – cap interactions. Binding of the cap analogues of the highest affinity is accompanied by less positive Cp due to the protein stiffening in the tightly bound complexes. The non-zero heat capacity changes cause that at biological temperatures recognition of the two natural dinucleotide cap analogues by eIF4E is both enthalpy- and entropy-driven, and stabilization of these complexes are relatively temperature-invariant. The exceptional, non-trivial and statistically important isothermal enthalpy-entropy compensation, with the characteristic temperature of 399 24 K, reveals important thermodynamic features of this macromolecular system: the dominating enthalpic character for the whole cap analogue series, a great instability of the apo-form of eIF4E, and significant stiffening of eIF4E in the cap-bound state. The presented results contribute to more profound insights into how eIF4E interacts with other components of the cytoplasmic machinery responsible for cap-dependent translation initiation. References 1. Varani, G. (1997) Structure.5, 855-858. 2. Furuichi, Y. and Shatkin, A. J. (2000) Advances in Virus Research, Vol 5555, 135-184. 3. Mattaj, I. W. (1986) Cell46, 905-911. 4. Hamm, J., Darzynkiewicz, E., Tahara, S. M., and Mattaj, I. W. (1990) Cell62, 569-577. 5. Sonenberg, N., Morgan, M. A., Merrick, W. C., and Shatkin, A. J. (1978) Proc. Natl. Acad. Sci. U. S. A75, 4843-4847. 6. Izaurralde, E., Lewis, J., McGuigan, C., Jankowska, M., Darzynkiewicz, E., and Mattaj, I. W. (1994) Cell78, 657-668. 7. Raught, B., Gingras, A. C., and Sonenberg, N. (2000) in Translational Control of Gene Expression pp 245-293, CSHL Press, New York. 8. De Benedetti, A. and Harris, A. L. (1999) Int. J. Biochem. Cell Biol.31, 59-72. 9. Marcotrigiano, J., Gingras, A. C., Sonenberg, N., and Burley, S. K. (1997) Cell89, 951-961. 10. Blachut-Okrasinska, E., Bojarska, E., Niedzwiecka, A., Chlebicka, L., Darzynkiewicz, E., Stolarski, R., Stepinski, J., and Antosiewicz, J. M. (2000) Eur. Biophys. J.29, 487-498. 11. Niedzwiecka, A., Marcotrigiano, J., Stepinski, J., Jankowska-Anyszka, M., Wyslouch-Cieszynska, A., Dadlez, M., Gingras, A. C., Mak, P., Darzynkiewicz, E., Sonenberg, N., Burley, S. K., and Stolarski, R. (2002) J. Mol. Biol.319, 615-635. 12. Niedzwiecka, A., Stepinski, J., Darzynkiewicz, E., Sonenberg, N., and Stolarski, R. (2002) Biochemistry41, 12140-12148. 13. Sharp, K. (2001) Protein Sci.10, 661-667. Acknowledgment Supported by the State Committee for Scientific Research 3P04A02125 and PBZ-KBN 059/T09/10