Download
1 / 1
10 likes | 126 Views
Effects of food and ectoparasites on dispersal age Victoria Garcia & Courtney J. Conway, Arizona Cooperative Fish and Wildlife Research Unit, School of Renewable Natural Resources, The University of Arizona, Tucson, AZ 85721. Results:.

E N D
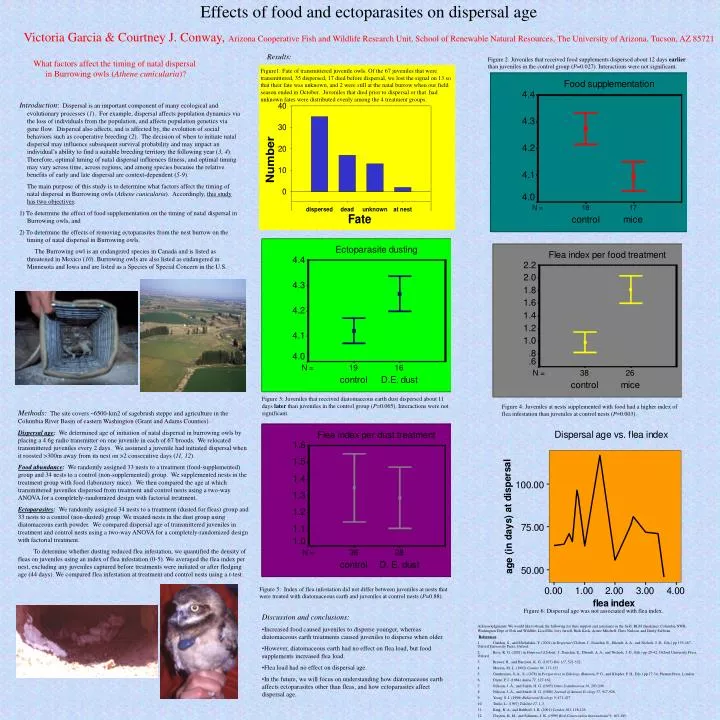
Effects of food and ectoparasites on dispersal age Victoria Garcia & Courtney J. Conway, Arizona Cooperative Fish and Wildlife Research Unit, School of Renewable Natural Resources, The University of Arizona, Tucson, AZ 85721 Results: Figure 2: Juveniles that received food supplements dispersed about 12 days earlier than juveniles in the control group (P=0.027). Interactions were not significant. What factors affect the timing of natal dispersal in Burrowing owls (Athene cunicularia)? Figure1: Fate of transmittered juvenile owls. Of the 67 juveniles that were transmittered, 35 dispersed, 17 died before dispersal, we lost the signal on 13 so that their fate was unknown, and 2 were still at the natal burrow when our field season ended in October. Juveniles that died prior to dispersal or that had unknown fates were distributed evenly among the 4 treatment groups. Introduction: Dispersal is an important component of many ecological and evolutionary processes (1). For example, dispersal affects population dynamics via the loss of individuals from the population, and affects population genetics via gene flow. Dispersal also affects, and is affected by, the evolution of social behaviors such as cooperative breeding (2). The decision of when to initiate natal dispersal may influence subsequent survival probability and may impact an individual’s ability to find a suitable breeding territory the following year (3, 4). Therefore, optimal timing of natal dispersal influences fitness, and optimal timing may vary across time, across regions, and among species because the relative benefits of early and late dispersal are context-dependent (5-9). The main purpose of this study is to determine what factors affect the timing of natal dispersal in Burrowing owls (Athene cunicularia). Accordingly, this study has two objectives: 1) To determine the effect of food supplementation on the timing of natal dispersal in Burrowing owls, and 2) To determine the effects of removing ectoparasites from the nest burrow on the timing of natal dispersal in Burrowing owls. The Burrowing owl is an endangered species in Canada and is listed as threatened in Mexico (10). Burrowing owls are also listed as endangered in Minnesota and Iowa and are listed as a Species of Special Concern in the U.S. Figure 3: Juveniles that received diatomaceous earth dust dispersed about 11 days later than juveniles in the control group (P=0.065). Interactions were not significant. Figure 4: Juveniles at nests supplemented with food had a higher index of flea infestation than juveniles at control nests (P=0.003). Methods:The site covers ~6500-km2 of sagebrush steppe and agriculture in the Columbia River Basin of eastern Washington (Grant and Adams Counties). Dispersal age: We determined age of initiation of natal dispersal in burrowing owls by placing a 4.6g radio transmitter on one juvenile in each of 67 broods. We relocated transmittered juveniles every 2 days. We assumed a juvenile had initiated dispersal when it roosted >300m away from its nest on >2 consecutive days (11, 12). Food abundance:We randomly assigned 33 nests to a treatment (food-supplemented) group and 34 nests to a control (non-supplemented) group. We supplemented nests in the treatment group with food (laboratory mice). We then compared the age at which transmittered juveniles dispersed from treatment and control nests using a two-way ANOVA for a completely-randomized design with factorial treatment. Ectoparasites:We randomly assigned 34 nests to a treatment (dusted for fleas) group and 33 nests to a control (non-dusted) group. We treated nests in the dust group using diatomaceous earth powder. We compared dispersal age of transmittered juveniles in treatment and control nests using a two-way ANOVA for a completely-randomized design with factorial treatment. To determine whether dusting reduced flea infestation, we quantified the density of fleas on juveniles using an index of flea infestation (0-5). We averaged the flea index per nest, excluding any juveniles captured before treatments were initiated or after fledging age (44 days). We compared flea infestation at treatment and control nests using a t-test. Figure 5: Index of flea infestation did not differ between juveniles at nests that were treated with diatomaceous earth and juveniles at control nests (P=0.88). Figure 6: Dispersal age was not associated with flea index. • Discussion and conclusions: • Increased food caused juveniles to disperse younger, whereas diatomaceous earth treatments caused juveniles to disperse when older. • However, diatomaceous earth had no effect on flea load, but food supplements increased flea load. • Flea load had no effect on dispersal age. • In the future, we will focus on understanding how diatomaceous earth affects ectoparasites other than fleas, and how ectoparasites affect dispersal age. Acknowledgments: We would like to thank the following for their support and assistance in the field: BLM (Spokane), Columbia NWR, Washington Dept of Fish and Wildlife, Lisa Ellis, Joey Jarrell, Rick Keck, Aimee Mitchell, Chris Nadeau, and Emily Sullivan. References 1. Gandon, S., and Michalakis, Y. (2001) in Dispersal (Clobert, J., Danchin, E., Dhondt, A. A., and Nichols, J. D., Eds.) pp 155-167, Oxford University Press, Oxford. 2. Ross, K. G. (2001) in Dispersal (Clobert, J., Danchin, E., Dhondt, A. A., and Nichols, J. D., Eds.) pp 29-42, Oxford University Press, Oxford. 3. Brewer, R., and Harrison, K. G. (1975) Ibis117, 521-522. 4. Morton, M. L. (1992) Condor94, 117-133. 5. Gauthreaux, S. A., Jr. (1978) in Perspectives in Ethology (Bateson, P. G., and Klopfer, P. H., Eds.) pp 17-54, Plenum Press, London. 6. Drent, P. J. (1984) Ardea72, 127-162. 7. Nilsson, J.-Å., and Smith, H. G. (1985) Ornis Scandinavica16, 293-298. 8. Nilsson, J.-Å., and Smith, H. G. (1988) Journal of Animal Ecology57, 917-928. 9. Yoerg, S. I. (1998) Behavioral Ecology9, 471-477. 10. Trulio, L. (1997) Tideline17, 1-3. 11. King, R. A., and Belthoff, J. R. (2001) Condor103, 118-126. 12. Clayton, K. M., and Schmutz, J. K. (1999) Bird Conservation International9, 163-185.






















