Download
1 / 1
10 likes | 214 Views
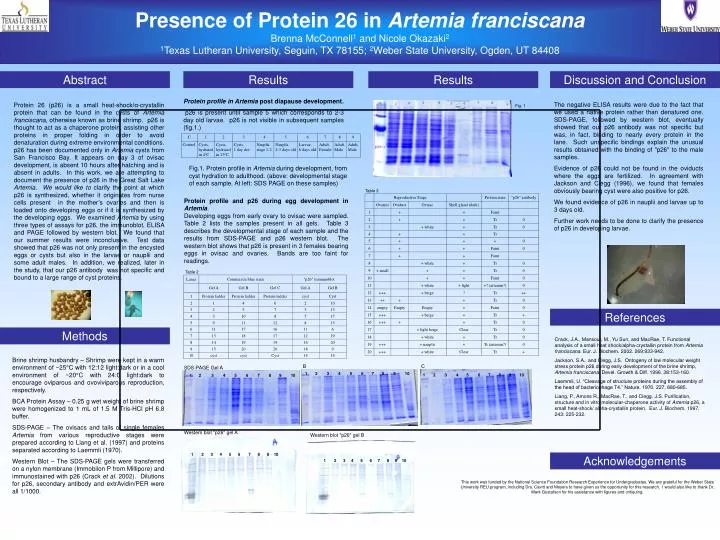
Presence of Protein 26 in Artemia franciscana Brenna McConnell 1 and Nicole Okazaki 2 1 Texas Lutheran University, Seguin, TX 78155; 2 Weber State University, Ogden, UT 84408. Fig. 1. 1 2 3 4 5 6 7 8 9 10.

E N D
Presence of Protein 26 in Artemia franciscana Brenna McConnell1 and Nicole Okazaki2 1Texas Lutheran University, Seguin, TX 78155; 2Weber State University, Ogden, UT 84408 Fig. 1 1 2 3 4 5 6 7 8 9 10 1 2 3 4 5 6 7 8 9 10 SDS-PAGE Gel A B C 1 2 3 4 5 6 7 8 9 10 Western blot "p26" gel A Western blot "p26" gel B 1 2 3 4 5 6 7 8 9 10 1 2 3 4 5 6 7 8 9 10 Abstract Results Results Discussion and Conclusion Protein profile in Artemia post diapause development. p26 is present until sample 5 which corresponds to 2-3 day old larvae. p26 is not visible in subsequent samples (fig.1.) Protein 26 (p26) is a small heat-shock/α-crystallin protein that can be found in the cysts of Artemia franciscana, otherwise known as brine shrimp. p26 is thought to act as a chaperone protein, assisting other proteins in proper folding in order to avoid denaturation during extreme environmental conditions. p26 has been documented only in Artemia cysts from San Francisco Bay. It appears on day 3 of ovisac development, is absent 10 hours after hatching and is absent in adults. In this work, we are attempting to document the presence of p26 in the Great Salt Lake Artemia. We would like to clarify the point at which p26 is synthesized, whether it originates from nurse cells present in the mother’s ovaries and then is loaded onto developing eggs or if it is synthesized by the developing eggs. We examined Artemia by using three types of assays for p26, the immunoblot, ELISA and PAGE followed by western blot. We found that our summer results were inconclusive. Test data showed that p26 was not only present in the encysted eggs or cysts but also in the larvae or nauplii and some adult males. In addition, we realized, later in the study, that our p26 antibody was not specific and bound to a large range of cyst proteins. The negative ELISA results were due to the fact that we used a native protein rather than denatured one. SDS-PAGE, followed by western blot, eventually showed that our p26 antibody was not specific but was, in fact, binding to nearly every protein in the lane. Such unspecific bindings explain the unusual results obtained with the binding of "p26" to the male samples. Evidence of p26 could not be found in the oviducts where the eggs are fertilized. In agreement with Jackson and Clegg (1996), we found that females obviously bearing cyst were also positive for p26. We found evidence of p26 in nauplii and larvae up to 3 days old. Further work needs to be done to clarify the presence of p26 in developing larvae. Fig.1. Protein profile in Artemia during development, from cyst hydration to adulthood. (above: developmental stage of each sample. At left: SDS PAGE on these samples) Table 3 Protein profile and p26 during egg development in Artemia. Developing eggs from early ovary to ovisac were sampled. Table 2 lists the samples present in all gels. Table 3 describes the developmental stage of each sample and the results from SDS-PAGE and p26 western blot. The western blot shows that p26 is present in 3 females bearing eggs in ovisac and ovaries. Bands are too faint for readings. Table 2 References Methods Crack, J.A., Mansour, M., Yu Sun, and MacRae, T. Functional analysis of a small heat shock/alpha-crystallin protein from Artemia franciscana. Eur. J. Biochem. 2002. 269:933-942. Jackson, S.A., and Clegg, J.S. Ontogeny of low molecular weight stress protein p26 during early development of the brine shrimp, Artemia franciscana. Devel. Growth & Diff. 1996. 38:153-160. Laemmli, U. “Cleavage of structure proteins during the assembly of the head of bacteriophage T4.” Nature. 1970. 227, 680-685. Liang, P., Amons R., MacRae, T., and Clegg, J.S. Purification, structure and in vitro molecular-chaperone activity of Artemia p26, a small heat-shock/ alpha-crystallin protein. Eur. J. Biochem. 1997, 243: 225-232. Brine shrimp husbandry – Shrimp were kept in a warm environment of ~25°C with 12:12 light:dark or in a cool environment of ~20°C with 24:0 light:dark to encourage oviparous and ovoviviparous reproduction, respectively. BCA Protein Assay – 0.25 g wet weight of brine shrimp were homogenized to 1 mL of 1.5 M Tris-HCl pH 6.8 buffer. SDS-PAGE – The ovisacs and tails of single females Artemia from various reproductive stages were prepared according to Liang et al. (1997) and proteins separated according to Laemmli (1970). Western Blot – The SDS-PAGE gels were transferred on a nylon membrane (Immobilon P from Millipore) and immunostained with p26 (Crack et al. 2002). Dilutions for p26, secondary antibody and extrAvidin/PER were all 1/1000. Acknowledgements This work was funded by the National Science Foundation Research Experience for Undergraduates. We are grateful for the Weber State University REU program, including Drs. Cavitt and Meyers to have given us the opportunity for this research. I would also like to thank Dr. Mark Gustafson for his assistance with figures and critiquing.