Download
1 / 104
1.04k likes | 1.04k Views
Nucleic acid structure informs function. We will revisit several aspects of nucleic acid structure that help us understand how DNA performs its functions. Nucleic Acid Structure. Andy Howard Macromolecular Structure 23 October 2018. Bases Nucleotides Polynucleotides Density Stability

E N D
Nucleic acid structure informs function • We will revisit several aspects of nucleic acid structure that help us understand how DNA performs its functions. Nucleic Acid Structure
Nucleic Acid Structure Andy HowardMacromolecular Structure23 October 2018
Bases Nucleotides Polynucleotides Density Stability Denaturation and renaturation A, B, Z DNA Tertiary structure Supercoiling Gyrases Nucleosomes Higher levels RNA Types & functions Bases & structures What we’ll discuss Nucleic Acid Structure
Pyrimidines • Single-ring nucleic acid bases • 6-atom ring; always two nitrogens in the ring,meta to one another • Based on pyrimidine, although pyrimidineitself is not a biologically important molecule • Variations depend on oxygens and nitrogens attached to ring carbons • Tautomerization possible • Note line of symmetry in pyrimidine structure 6 5 1 4 2 3 Nucleic Acid Structure
Uracil and thymine • Uracil is a simple dioxo derivative of pyrimidine: 2,4-dioxopyrimidine • Thymine is 5-methyluracil • Uracil is found in RNA; Thymine is found in DNA • We can draw other tautomers where we move the protons to the oxygens Nucleic Acid Structure
Tautomers • Lactam and lactim forms • Getting these right was essential to Watson & Crick’s development of the DNA double helical model Nucleic Acid Structure
Cytosine • This is 2-oxo,4-aminopyrimidine • It’s the other pyrimidine basefound in DNA & RNA • Can undergo spontaneous deamination (CU) • Again, other tautomers can be drawn Nucleic Acid Structure
Cytosine:amino and imino forms • Again, this tautomerization needs to be kept in mind Nucleic Acid Structure
7 6 Purines 5 1 8 4 2 9 3 • Derivatives of purine; again, the root molecule isn’t biologically important • Six-membered ring looks a lot like pyrimidine • Numbering works somewhat differently: note that the glycosidic bonds will be to N9, whereas it’s to N1 in pyrimidines Nucleic Acid Structure
Adenine • This is 6-aminopurine • Found in RNA and DNA • We’ve seen how important adenosine and its derivatives are in metabolism • Tautomerization happens here too Nucleic Acid Structure
Guanine • This is 2-amino-6-oxopurine • Found in RNA, DNA • Lactam, lactim forms Nucleic Acid Structure
Other natural purines • Hypoxanthine and xanthine are biosynthetic precursors of A & G • Urate is important in nitrogen excretion pathways Nucleic Acid Structure
Tautomerization and H-bonds • Lactam forms predominate at neutral pH • This influences which bases are H-bond donors or acceptors • Amino groups in C, A, G make H-bonds • So do ring nitrogens at 3 in pyrimidines and 1 in purines • … and oxygens at 4 in U,T, 2 in C, 6 in G Nucleic Acid Structure
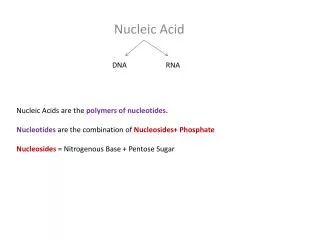
Nucleosides • These are glycosides (covalent adducts containing sugar) of the nucleic acid bases • Sugar is always ribose or deoxyribose, in their furanose (5-membered ring) form • Connected nitrogen is: • N1 for pyrimidines (on 6-membered ring) • N9 for purines (on 5-membered ring) Nucleic Acid Structure
Pyrimidine nucleosides • Drawn here in amino and lactam forms uridine cytidine Nucleic Acid Structure
Pyrimidine deoxynucleosides deoxyuridine deoxythymidine deoxycytidine Nucleic Acid Structure
A tricky nomenclature issue • Remember that thymidine and its phosphorylated derivatives ordinarily occur associated with deoxyribose, not ribose • Therefore many people leave off the deoxy- prefix in names of thymidine and its derivatives: it’s usually assumed. Nucleic Acid Structure
Purine nucleosides • Drawn in amino and lactam forms adenosine guanosine Nucleic Acid Structure
Purine deoxynucleosides deoxyadenosine deoxyguanosine Nucleic Acid Structure
Monophosphorylated nucleosides • We have specialized names for the 5’-phospho derivatives of the nucleosides, i.e. the nucleoside monophosphates: • They are nucleotides, including Adenosine 5’-monophosphate = AMP = adenylate adenylate Nucleic Acid Structure
Three other nucleotides cytidylate = CMP guanylate = GMP uridylate = UMP Nucleic Acid Structure
Deoxynucleotides • Similar nomenclature • dUMP = deoxyuridylate • dAMP = deoxyadenylate • dGMP = deoxyguanylate • dCMP = deoxycytidylate • dTMP (= TMP) = deoxythymidylate = thymidylate Deoxy-uridylate = dUMP Nucleic Acid Structure
Di- and triphosphates • Phosphoanhydride bonds link second and perhaps third phosphates to the5’-OH on the ribose moiety CTP GDP Nucleic Acid Structure
Cyclic phosphodiesters • 3’ and 5’ hydroxyls are both involved in -O-P-O bonds • cAMP and cGMP are the important ones Cyclic AMP Nucleic Acid Structure
Glycosidic bonds • This illustrates the roughly perpendicular positionings of the base and sugar rings Nucleic Acid Structure
Conformations around the glycosidic bond • Rotation of the base around the glycosidic bond is sterically hindered • In the syn conformation there would be some interference between the sugar ring and the 2-position of the base Nucleic Acid Structure
How this really plays out • Therefore pyrimidines are always anti, and purines are usually anti • Furanose and base rings are roughly perpendicular Nucleic Acid Structure
Nucleotide Oligomers and Polymers • Monomers are nucleotides or deoxynucleotides • Linkages are phosphodiester linkages between 3’ of one ribose and 5’ of the next ribose • It’s logical to start from the 5’ end for synthetic reasons Nucleic Acid Structure
Typical DNA dinucleotide • Various notations: this is pdApdCp • Leave out the p’s if there’s a lot of them! Nucleic Acid Structure
DNA structure • Many years of careful experimental work enabled fabrication of double-helical model of double-stranded DNA • Explained [A]=[T], [C]=[G] Nucleic Acid Structure
Stability of the double helix • Specific H-bonds stabilize double-helical structure • These H-bonds aren’t responsible for all of the stability of the double helix! Nucleic Acid Structure
What does double-stranded DNA really look like? • Picture on earlier slide emphasizes only the H-bond interactions; it ignores the orientation of the sugars, which are actually tilted relative to the helix axis • Planes of the bases are almost perpendicular to the helical axes on both sides of the double helix Nucleic Acid Structure
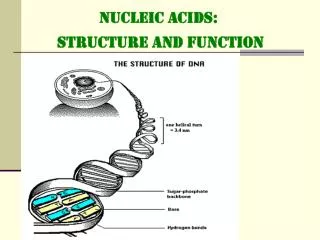
Sizes • Diameter of the double helix: 2.37nm • Length along one full turn:10.4 base pairs = pitch = 3.40nm • Distance between stacked base pairs = rise = 0.33 nm • Major groove is wider and shallower;minor groove is narrower and deeper Nucleic Acid Structure
GC-rich DNA is denser • DNA is denser than RNA or protein, period, because it can coil up so compactly • Therefore density-gradient centrifugation separates DNA from other cellular macromolecules • GC-rich DNA is 3% denser than AT-rich • Can be used as a quick measure of GC content Nucleic Acid Structure
Density as function of GC content Nucleic Acid Structure
Base composition for DNA • As noted, [A]=[T], [C]=[G] because of base pairing • [A]/[C] etc. not governed by base pairing • Can vary considerably • E.coli : [A], [C] about equal • Mycobacterium tuberculosis: [C] > 2*[A] • Mammals: [C] < 0.74*[A] Nucleic Acid Structure
What stabilizes DNA? • Variety of stabilizing interactions • Stacking of base pairs • Hydrogen bonding between base pairs • Hydrophobic effects (burying bases, which are less polar) • Charge-charge interactions:phosphates with Mg2+ & cationic proteins Courtesy dnareplication.info Nucleic Acid Structure
How close to instability is it? • Pretty close. • Heating DNA makes it melt • pH > 10 separates strands too • The more GC pairs, the harder it is to melt DNA thermally • Weaker stacking interactions in A-T • One more H-bond per GC than per AT Nucleic Acid Structure
Denaturing and Renaturing DNA • When DNA is heated to > 80ºC, its UV absorbance increases by 30-40% • This hyperchromic shift reflects the unwinding of the DNA double helix • Stacked base pairs in native DNA absorb less light • When T is lowered, the absorbance drops, reflecting the re-establishment of stacking Nucleic Acid Structure
Heat denaturation • Figure 11.14Heat denaturation of DNA from various sources, so-called melting curves. The midpoint of the melting curve is defined as the melting temperature, Tm.(From Marmur, J., 1959. Nature183:1427–1429.) Nucleic Acid Structure
GC content vs. melting temp • High salt and no chelators raises the melting temperature Nucleic Acid Structure
How else can we melt DNA? • High pH deprotonates the bases so the H-bonds disappear • Low pH hyper-protonates the bases so the H-bonds disappear • Alkalai is better: it doesn’t break the glycosidic linkages • Urea, formamide make better H-bonds than the DNA itself so they denature DNA Nucleic Acid Structure
What happens if we separate the strands? • We can renature the DNA into a double helix • Requires re-association of 2 strands: reannealing • The realignment can go wrong • Association is 2nd-order, zippering is first order and therefore faster Nucleic Acid Structure
Steps in denaturation and renaturation Nucleic Acid Structure
Rate depends on complexity • The more complex DNA is, the longer it takes for nucleation of renaturation to occur • “Complex” can mean “large”, but complexity is influenced by sequence randomness: poly(AT) is faster than a random sequence Nucleic Acid Structure
Second-order kinetics • Rate of association: -dc/dt = k2c2 • Boundary condition is fully denatured concentration c0 at time t=0: • c / c0 = (1+k2c0t)-1 • Half time is t1/2 = (k2c0)-1 • Routine depiction: plot c0t vs. fraction reassociated (c /c0) and find the halfway point. Nucleic Acid Structure
Typical c0t curves Nucleic Acid Structure
Helical twist • Rotation about the backbone axis • Successive base-pairs rotated with respect to each other by ~ 32º Nucleic Acid Structure