Download

1 / 28
300 likes | 852 Views
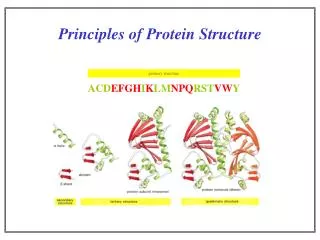
Principles of Protein Structure . PHAR 201/Bioinformatics I Philip E. Bourne School of Pharmacy & Pharm. Sci., UCSD Prerequisite Reading: Structural Bioinformatics Chapters 1-2 Thanks to Eric Scheeff and Lynn Fink. Remember .

E N D
Principles of Protein Structure PHAR 201/Bioinformatics I Philip E. Bourne School of Pharmacy & Pharm. Sci., UCSD Prerequisite Reading: Structural Bioinformatics Chapters 1-2 Thanks to Eric Scheeff and Lynn Fink PHAR201 Lecture 1 2012
Remember .. • The first 2 lectures are not so much to teach/refresh your knowledge of protein/DNA/RNA structure, but for you to conceptualize, describe and subsequently analyze complex biological data • Assignment 1 will test this PHAR201 Lecture 1 2012
Remember.. • All which we study is an abstraction to make comprehension of a complex entity more straightforward • We think of structures as static entities, but they are dynamic, sometimes to the point of being ill-definable – function requires this flexibility • The more we have the more we should know and use – contrast Kendrew to the PDB today PHAR201 Lecture 1 2012
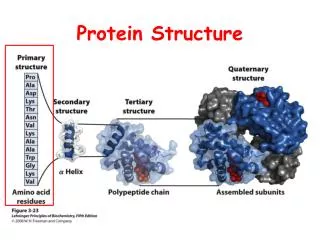
Primary Structure - Amino Acids • It is the amino acid sequence (1940) that “exclusively” determines the 3D structure of a protein • 20 amino acids – modifications do occur post translationally PHAR201 Lecture 1 2012
Amino Acids Continued… • It is the properties of the R group that determine the property of the aa and ultimately the protein • Different schemes exist for describing the properties Willie Taylor’s scheme is often employed in bioinformatics analyses • Hydrophobicity, polarity and charge are common measures • Learn the amino acid codes, structures and properties! PHAR201 Lecture 1 2012 Primary Structure
Amino Acids Continued… • Chirality – amino acids are enatiomorphs, that is mirror images exist – only the L(S) form is found in naturally forming proteins. Some enzymes can produce D(R) amino acids • Think about a data structure for this information – annotation and a validation procedure should be included • Think about systematic versus common nomenclature PHAR201 Lecture 1 2012 Primary Structure
Peptide Bond Formation • Individual amino acids form a polypeptide chain • Such a chain is a component of a hierarchy for describing macromolecular structure • The chain has its own set of attributes • The peptide linkage is planar and rigid PHAR201 Lecture 1 2012 Primary Structure
Geometry of the Chain • A dihedral angle is the angle between two planes defined by 4 atoms – 123 make one plane; 234 the other • Omega is the rotation around the peptide bond Cn – Nn+1 – it is planar and is 180 under ideal conditions • Phi is the angle around N – Calpha • Psi is the angle around Calpha C’ • The values of phi and psi are constrained to certain values based on steric clashes of the R group. Thus these values show characteristic patterns as defined by the Ramachandran plot PHAR201 Lecture 1 2012 From Brandon and Tooze Secondary Structure
Ramachandran Plot • Shows allowed and disallowed regions • Gly and Pro are exceptions: Gly has no limitation; Pro is constrained by the fact its side chain binds back to the main chain Gray = allowed conformations. βA, antiparallel b sheet; βP, parallel b sheet; βT, twisted b sheet (parallel or anti-parallel); α, right-handed α helix; L, left-handed helix; 3, 310 helix; p, Π helix. PHAR201 Lecture 1 2012 Secondary Structure
Secondary Structure • The chemical nature of the carboxyl and amino groups of all amino acids permit hydrogen bond formation (stability) and hence defines secondary structures within the protein. • The R group has an impact on the likelihood of secondary structure formation (proline is an extreme case) • This leads to a propensity for amino acids to exist in a particular secondary structure conformation • Helices and sheets are the regular secondary structures, but irregular secondary structures exist and can be critical for biological function PHAR201 Lecture 1 2012 Secondary Structure
Alpha Helix • A helix can turn right or left from N to C terminus – only right-handed are observed in nature as this produces less clashes • All hydrogen bonds are satisfied except at the ends = stable PHAR201 Lecture 1 2012 Secondary Structure
Alpha Helix Continued • There are 3.6 residues per turn • A helical wheel will outline the surface properties of the helix PHAR201 Lecture 1 2012 Secondary Structure
Other (Rarer) Helix Types - 310 • Less favorable geometry • 3 residues per turn with i+3 not i+4 • Hence narrower and more elongated • Usually seen at the end of an alpha helix PHAR201 Lecture 1 2012 4HHB Secondary Structure
Other (Very Rare) Helix Types - Π • Less favorable geometry • 4 residues per turn with i+5 not i+4 • Squat and constrained PHAR201 Lecture 1 2012 Secondary Structure
Beta Sheets PHAR201 Lecture 1 2012 Secondary Structure
Beta Sheets Continued • Between adjacent polypeptide chains • Phi and psi are rotated approximately 180 degrees from each other • Mixed sheets are less common • Viewed end on the sheet has a right handed twist that may fold back upon itself leading to a barrel shape (a beta barrel) • Beta bulge is a variant; residue on one strand forms two hydrogen bonds with residue on other – causes one strand to bulge – occurs most frequently in parallel sheets PHAR201 Lecture 1 2012 Secondary Structure
Other Secondary Structures – Loop or Coil • Often functionally significant • Different types • Hairpin loops (aka reverse turns) – often between anti-parallel beta strands • Omega loops – beginning and end close (6-16 residues) • Extended loops – more than 16 residues 1AKK PHAR201 Lecture 1 2012 Secondary Structure
Tertiary Structure • Myoglobin (Kendrew 1958) and hemoglobin (Perutz 1960) gave us the proven experimental insights into tertiary structure as secondary structures interacting by a variety of mechanisms • While backbone interactions define most of the secondary structure interactions, it is the side chains that define the tertiary interactions PHAR201 Lecture 1 2012 Tertiary Structure
Components of Tertiary Structure • Fold – used differently in different contexts – most broadly a reproducible and recognizable 3 dimensional arrangement • Domain – a compact and self folding component of the protein that usually represents a discreet structural and functional unit • Motif (aka supersecondary structure) a recognizable subcomponent of the fold – several motifs usually comprise a domain Like all fields these terms are not used strictly making capturing data that conforms to these terms all the more difficult PHAR201 Lecture 1 2012 Tertiary Structure
Tertiary Structure as Dictated by the Environment • Proteins exist in an aqueous environment where hydrophilic residues tend to group at the surface and hydrophobic residues form the core – but the backbone of all residues is somewhat hydrophilic – therefore it is important to have this neutralized by satisfying all hydrogen bonds as is achieved in the formation of secondary structures • Polar residues must be satisfied in the same way – on occasion pockets of water (discreet from the solvent) exist as an intrinsic part of the protein to satisfy this need • Ion pairs (aka salt bridge) form important interactions • Disulphide linkages between cysteines form the strongest (ie covalent tertiary linkages); the majority of cysteines do not form such linkages 5EBX PHAR201 Lecture 1 2012 Tertiary Structure
Tertiary Structure as Dictated by Protein Modification • To the amino acid itself eg hydroxyproline needed for collagen formation • Addition of carbohydrates (intracellular localization) • Addition of lipids (binding to the membrane) • Association with small molecules – notably metals eg hemoglobin PHAR201 Lecture 1 2012 Tertiary Structure
There are Different Forms of Classification apart from Structural • Biochemical • Globular • Membrane • Fibrous myoglobin Bacteriorhodopsin Collagen PHAR201 Lecture 1 2012
Quaternary Structure • The biological function of some molecules is determined by multiple polypeptide chains – multimeric proteins • Chains can be identical eg homeodimer or different eg heterodimer • The interactions within multimers is the same as that found in tertiary and secondary structures PHAR201 Lecture 1 2012
Hemoglobin: Enhanced binding capability of oxygen Cooperativity Glutamine sythetase: Controlled use of Nitrogen from Multiple active sites Co-location of Function Combination Immunoglobulin: Multiple receptor responses Structural Assembly Actin: Giving the cell shape and form PHAR201 Lecture 1 2012 Quaternary Structure
Quaternary Structure: Ferritin - The Bodies Iron Storage Protein PHAR201 Lecture 1 2012 Quaternary Structure
Disorder? PHAR201 Lecture 1 2012
Additional Reading • Branden and Tooze (1999) Introduction to Protein Structure (2nd Edition) Garland Publishing. An excellent introduction • Richardson (1981) The Anatomy and Taxonomy of Protein Structure Adv. Protein Chem. 34: 167-339 Good historical perspective PHAR201 Lecture 1 2012