Download
1 / 66
660 likes | 886 Views
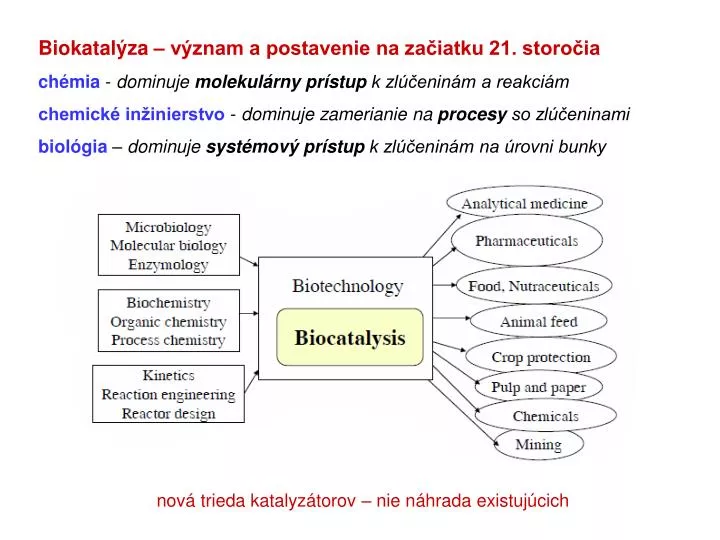
Biokatalýza – význam a postavenie na začiatku 21. storočia chémia - dominuje molekulárny prístup k zlúčeninám a reakciám chemické inžinierstvo - dominuje zamerianie na procesy so zlúčeninami biológia – dominuje systémový prístup k zlúčeninám na úrovni bunky.

E N D
Biokatalýza – význam a postavenie na začiatku 21. storočia chémia - dominuje molekulárny prístup k zlúčeninám a reakciám chemické inžinierstvo - dominuje zamerianie na procesy so zlúčeninami biológia – dominuje systémový prístup k zlúčeninám na úrovni bunky nová trieda katalyzátorov – nie náhrada existujúcich
Enzým je makromolekula, obvykle proteín často obsahujúci alebo vyžadujúci jeden alebo viac kovových iónov, ktorá pôsobí ako (bio)katalyzátor zvýšením reakčného toku. Enzým všeobecne katalyzuje len jeden typ reakcie (reakčná selektivita) a pôsobí len na obmedzenú množinu substrátov, alebo len na jeden substrát (substrátová selektivita). Molekuly substrátu sa transformujú na rovnakej funkčnej skupine (regioselektivita), pričom sa transformuje len jeden substrát alebo jeden z enantiomerov, chirálnych substrátov alebo racemických zmesí (enantioselektivita)
Kofaktor (cofactor) je organická molekula alebo ión (obvykle ión kovu) nutný pre aktivitu enzýmu. Voľne pripojený sa nazýva koenzým, kovalentne pripojený kofaktor sa nazýva prostetická skupina Koenzým(coenzyme) je disociovateľná nízkomolekulárna neproteínová organická látka (obvykle nukleotid), ktorá se zúčastňuje enzýmových reakcií ako akceptor nebo donor chemických skupín alebo elektrónov. Prostetická skupina(prostethic group) je v enzýme pevne viazaná špecifická nepeptidová zlúčenina zúčastňujúca sa jeho biologickej aktivity Väzbové miesto (binding site) je špecifická oblasť (alebo atóm) molekuly enzýmu, ktoré je schopné vstúpiť do stabilizujúcej interakcie s inou molekulou. Aktívne miesto (active site, active center) je oblasť enzýmu, kde prebieha špecifická reakcia.
Stickase Substrate Transition state Product If enzyme just binds substrate then there will be no further reaction X Enzyme not only recognizes substrate, but also induces the formation of transition state Adapted from Nelson & Cox (2000) Lehninger Principles of Biochemistry (3e) p.252
B A The Nature of Enzyme Catalysis ●Enzyme provides a catalytic surface ●This surface stabilizes transition state ● Transformed transition state to product B A Catalytic surface
Active Site Is a Deep Buried Pocket Why energy required to reach transition state is lower in the active site? It is a magic pocket (1) Stabilizes transition + (2) Expels water CoE (2) (1) (3) Reactive groups (4) - (4) Coenzyme helps (3)
Enzyme Active Site Is Deeper than Ab Binding Instead, active site on enzyme also recognizes substrate, but actually complementally fits the transition state and stabilized it. Ag binding site on Ab binds to Ag complementally, no further reaction occurs. X Adapted from Nelson & Cox (2000) Lehninger Principles of Biochemistry (3e) p.252
Active Site Avoids the Influence of Water + - Preventing the influence of water sustains the formation of stable ionic bonds Adapted from Alberts et al (2002) Molecular Biology of the Cell (4e) p.115
Príklady enzýmových reakcií so syntetickými organickými zlúčeninami
Priama a nepriama epoxidácia katalyzovaná haloperoxidázami
Enzyme Kinetics Direct plot Significance kcat/Km zero order 1st order Vmax[S] kcat vo= E3 Km+[S] Turn over number E2 E1 k3 [Et] Competitive Double reciprocal Activity Unit 1 mmole min Non-competitive Specific Activity unit mg Uncompetitive Observe vo change under various [S], resulted plots yield Vmax and Km Vmax Km & Maximum velocity Affinity with substrate Inhibition Bi-substrate reaction also follows M-M equation, but one of the substrate should be saturated when estimate the other Activity Juang RH (2004) BCbasics
Significance of Enzyme Kinetics Vmax[S] vo = Km+[S] v0 = Vmax × K = k3 [Et]× K ObtainVmaxandKm zero order E3 Proportional to enzyme concentration E2 E1 1st order [S] = Low → High [S] = Fixed concentration Juang RH (2004) BCbasics
Enzyme Activity Unit Specific Activity y x = = tan S →Pmmole t Product [P] vo = [P]/min Slope tan mmole /min Unit = 0 10 20 30 40 Reaction time(min) Activity Units y Juang RH (2004) BCbasics Protein (mg) x
Increase Substrate Concentration 0 1 2 3 4 5 6 7 8 80 60 40 20 0 Product 0 2 4 6 8 Substrate (mmole) S + E ↓ P (in a fixed period of time) Juang RH (2004) BCbasics
An Example for Enzyme Kinetics (Invertase) 1 vo vo 1 Vmax - 1 Km 1/S S 1)Use predefined amount ofEnzyme→E 2)Add substratein various concentrations →S(x) 3)MeasureProductin fixedTime (P/t) →vo(y) 4)(x, y)plot get hyperbolic curve,estimate→Vmax 5)Wheny = 1/2 Vmaxcalculatex ([S]) →Km Vmax 1/2 Juang RH (2004) BCbasics Km Double reciprocal Direct plot
A Real Example for Enzyme Kinetics Substrate Product Velocity Double reciprocal 2.0 1.0 0 1.0 0.5 0 v 1/v 1.0 -3.8 -4 -2 0 2 4 0 1 2 [S] 1/[S] Data v (mmole/min) [S] Absorbance 1/S 1/v no 0.25 0.50 1.0 2.0 0.21 0.36 0.40 0.46 0.42 0.72 0.80 0.92 4 2 1 0.5 2.08 1.56 1.35 1.16 1 2 3 4 → → → → (1) The product was measured by spectroscopy at 600 nm for 0.05 per mmole (2) Reaction time was 10 min Direct plot Double reciprocal Juang RH (2004) BCbasics
Enzyme Inhibition (Mechanism) E + S→ES→E + P + I ↓ EI E + S→ES→E + P + + II ↓ ↓ EI+S→EIS E + S→ES→E + P + I ↓ EIS ← ← ← ↑ ↑ ↑ ↑ Uncompetitive Competitive Non-competitive E Substrate E X Cartoon Guide Compete for active site Inhibitor Different site Equation and Description [I] binds to free [E] only, and competes with [S]; increasing [S] overcomes Inhibition by [I]. [I] binds to [ES] complex only, increasing [S] favors the inhibition by [I]. [I] binds to free [E] or [ES] complex; Increasing [S] can not overcome [I] inhibition. Juang RH (2004) BCbasics
Enzyme Inhibition (Plots) Uncompetitive Competitive Non-competitive Vmax Vmax vo Vmax’ Vmax’ I Direct Plots Km [S], mM Km’ Km [S], mM 1/vo 1/vo 1/vo I I Double Reciprocal Two parallel lines Intersect at X axis Intersect at Y axis 1/Vmax 1/Vmax 1/Vmax 1/Km 1/[S] 1/Km 1/[S] 1/Km 1/[S] Vmax vo I I Km Km’ [S], mM =Km’ Vmax unchanged Km increased Vmax decreased Km unchanged Both Vmax & Km decreased I Juang RH (2004) BCbasics
Porovnanie rýchlostných konštánt niektorých enzýmov v log škále ADC: arginine decarboxylase ODC: orotidine 5′-phosphate decarboxylase; STN: staphylococcal nuclease; GLU: sweet potato β-amylase; FUM: fumarase; MAN: mandelate racemase; PEP: carboxypeptidase B; CDA: E. coli cytidine deaminase; KSI: ketosteroid isomerase; CMU: chorismate mutase; CAN: carbonic anhydrase.
Niekoľko príkladov moderných aplikácií enzýmov
Mikročipy malých molekúl (small-molecule microarrays) (a)detekčná stratégia pomocou hydrolytických enzýmov (b)stratégia screeningu inhibítorov v roztoku CurrentOpinioninChemicalBiology2005,9:4–13
Stratégia pre špecifické chemické značenie proteínov in vivo (a) transfekcia živých buniek s DNA kódujúcou sledovaný proteín (POI - protein ofinterest), ktorý je sfúzovaný s proteínovým alebo peptidovým receptorom (b) pridanie sondy s detekovateľ-nou značkou (Tag) k bunkám po expresiifúzovaného proteínu;sonda (probe)je malá molekula, priepustná pre bunky, obsahujúca ligand s pripojenou značkou, ktorá sa pridá do kultivačného média (c) analýza funkcie proteínu v bunke pomocou fluorescenčnej mikroskopie alebo inej detekcie Current Opinion in Chemical Biology 2005, 9:56–61
Metódy chemického značenia fúzovaných proteínov in vivo Fluorescenčné značenie proteínov bisarzenitom FlAsH 4',5'-bis(1,3,2-dithioarsolan-2-yl)fluorescein Bisarzenitový derivát fluoresceínu sa viaže so subnanomolárnou afinitou na krátky tetracysteínový peptid fúzovaný so sledovaným proteínom POI Current Opinion in Chemical Biology 2005, 9:56–61
Metódy chemického značenia fúzovaných proteínov in vivo Kovalentné značenie ľudskej O6-alkylguanín-DNA-alkyltransferázy (hAGT) pomocou benzylguanínových derivátov Current Opinion in Chemical Biology 2005, 9:56–61
Metódy chemického značenia fúzovaných proteínov in vivo Schéma fúzneho proteínu s E. coli dihydrofolát reduktáznou doménou (eDHFR), do ktorej je naviazaný fluorescenčný konjugát metotrexát-Texas RedTM Current Opinion in Chemical Biology 2005, 9:56–61
Metódy chemického značenia fúzovaných proteínov in vivo Značenie pomocou splicingu proteínov (split intein) N-koncová časť inteínu sa exprimuje ako zmesný proteín s POI. C-koncová časť inteínu sa kovalentne označí chemickou značkou. Po kombinácii častí inteínu a následnom self-splicingu vznikne značený POI. Current Opinion in Chemical Biology 2005, 9:56–61






















