Download

1 / 75
770 likes | 1.11k Views
The Cell Cycle and its implications in diseases. Hansjörg Hauser Dept. of Gene Regulation and Differentiation Molecular Biotechnology HZI, Braunschweig. Cell division is a prerequisite for life. Microorganisms reproduce by cell division

E N D
The Cell Cycle and its implications in diseases Hansjörg Hauser Dept. of Gene Regulation and Differentiation Molecular Biotechnology HZI, Braunschweig
Cell division is a prerequisite for life • Microorganisms reproduce by cell division • Mammals need cell division during embryogenesis and for tissue homeostasis Example: Adult humans produce several milions of new cells per second (more than 1011 per day – about 100 grams)
Cell division can be fast or slow • Microorganisms > 20 min per division • Multicellular organisms: 8 min and several weeks per division • All species can halt cell division
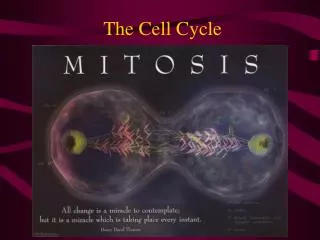
Neben einem zum Vergleich dargestellten Zellkern in der Interphase sind verschiedene Stadien der Mitose gezeigt (entsprechend der deutschen Literatur, daher ohne Prometaphase).
Schematische Darstellung des Zellzyklus. Zur besseren Veranschaulichung sind Chromosomen hier auch in Interphase so gezeichnet, wie sie in der Mitose aussehen. Dies entspricht jedoch nicht der Wirklichkeit.
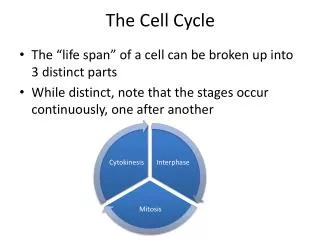
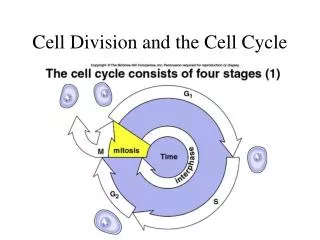
The Cell Cycle Schematic of the cell cycle. Outer ring: I=Interphase, M=Mitosis; Inner ring: M=Mitosis, G1=Gap 1, G2=Gap 2, S=Synthesis; Not in ring: G0=Gap 0/Resting.
Cell cycle control was studied in early embryogenesis of frogs, yeasts and mammalian cells. The mechanisms and involved molecules are highly conserved The names are sometimes confusing!
Cell division and duplication of cellular constituents • DNA • Proteins, Lipids, Carbohydrates • Organelles, Membranes...
Standard versus early embryonic cell cycle • in early embryonic cells DNA replication is uncoupled from other synthesis • The eggs contain more cytoplasm than normal cells: Stocks • Some organisms have eggs with 100.000times more cytoplasm than normal cells • 16 –17 divisions are possible without significant protein production. • These divisions are running without feedback control
Methods to measure cell division • Counting • Amount of DNA • Enzymatic activities • Incorporation of labeled DNA precursors • Cell cycle analysis (FACS) • Dilution of dyes (CFSE) • Time lapse microscopy
CFSE (Carboxyfluorescein succinimidyl ester) is a fluorescent cell staining dye. CFSE is commonly confused with CFDA-SE, although they are not strictly the same molecule; CFDA-SE is cell permeable, while CFSE is not. As CFDA-SE enters the cell cytoplasm, intracellular esterases convert the molecule to the fluorescent ester, CFSE, which is retained within cells. CFSE is a simple and sensitive technique for analysis of multiple parameters of cells. This method allows us to examine specific populations of proliferating cells and identify 7–10 successive cell generations, which has first been employed to detect proliferation of T cells in experimental animals.[1] CFSE consists of a fluorescent molecule containing a succinimydyl ester functional group and two acetate moieties. CFSE diffuses freely inside the cells and intracellular esterases cleave the acetate groups converting it to a fluorescent, membrane impermeable dye. This dye is not transferred to adjacent cells. CFSE is retained by the cell in the cytoplasm and does not adversely affect cellular function. During each round of cell division, relative fluorescence intensity of the dye is decreased by half. In addition, unlike other methods, CFSE-labeled viable cells can be recovered for further analysis.
FACS profiles of resting and growing cells G1 82% S 12% G2 6% G1 33% S 47% G2 19%
Kinases control the progression through the cell cycle Example MPF (maturation promoting factor) MPF is composed of a cyclin and a cyclin dependent protein kinase (cdk)
While cdks are constitutively expressed the appearance of cyclins in the cell cycle is transient – they cycle The presence of cyclins regulates the activity of the cdks
Cyclic activity of Cyclin kinases Temporal control of the animal cell cycle. The cyclin-E-, cyclin-A- and cyclin-B-dependent kinases are active at different times in the cell cycle. On this basis, cyclin E–Cdk2 appears to have a role in promoting S phase, cyclin A–Cdk2 in S phase and at G2-to-M phase, and cyclin B–Cdk1 during mitosis. Cyclin B1–Cdk1 is activated at the end of G2 phase by the phosphatase Cdc25.
Targets of cyclin kinases: the G2 Kinase complex (MPF)
The kinase activity of cdc-cyclin compexes is regulated by phosphorylation and dephosphorylation Example MO16 is an activating kinase Wee1 is an inhibitory kinase cdc25 is a phosphatase that removes the inhibitory phosphate from the cdk
Regulation of cyclin-dependent kinases. Arrowheads represent activating events and perpendicular ends represent inhibitoryevents. Genes known to perform the indicated functions are listedbelow. Both cyclins and some CKIs (Cdk inhibitors) are regulatedby synthesis and ubiquitin-mediated proteolysis. Checkpoint pathwayscould act to promote inhibitory pathways or inhibit activatingpathways to cause cell cycle arrest
The progression through the cell cycle underlies many controls: Example DNA replication A re-replication block ensures that no segment of DNA is replicated more than once Passage through mitosis removes the re-replication block Feedback controls generally depend on inhibitory signals
Checkpoint pathways (A) A genetic pathway illustrating intrinsic and extrinsic checkpoint mechanisms. Letters representcell cycle processes. The pathway shown as red symbols indicates an intrinsiccheckpoint mechanism that operates to ensure that event C is completedbefore event E. After event B is completed, an inhibitory signalis activated that blocks completion of event E. After event Cis completed, a signal is sent to turn off the inhibitory signalfrom B, thereby allowing completion of E. The blue symbols representan extrinsic mechanism that is activated when defects such asDNA damage or spindle errors are detected. It is arbitrarily locatedon the D to E pathway but could also function by inhibiting alater step in the B to C pathway. In that case, the extrinsicpathway would utilize the intrinsic mechanism for cell cycle arrest.Mutations in any of the red or blue symbols would result in acheckpoint-effective phenotype.
Checkpoint pathways (B) Schematic representationof several cell cycle checkpoints. The colored arrows depict complexsignaling pathways that operate in G1 to transmit informationregarding cell proliferation. The red lines connecting particularevents and cell cycle transitions represent the inhibitory signalsgenerated by checkpoint pathways in response to those events.The points of contact of the negative growth factor and contactinhibition pathways with the cell cycle are arbitrary and meantto indicate arrest in G1.
Activation of DNA replication through gene expression in G1 G1 S G0 Activation of delayed response genes: E2F Cyclins E, D DNA synthesis genes early response genes: fos, jun,..
Resting cells: The retinoblastoma protein Rb blocks cell cycle progression in G1 by binding to and sequestering E2F Rb-P Rb + E2F Phosphorylation causes Inactivation of Rb Rb : E2F
Cell cycle progression by growth factors Phosphorylation causes Inactivation of Rb Proliferation Rb-P Rb E2F CyclinD.cdk4 Rb: E2F CyclinD Rb captures E2F: E2F cannot activate proproliferative genes MAPK pathway Ras EGF
Cell cycle progression Proliferation Growth block Phosphorylation causes Inactivation of Rb Rb captures E2F, so that it cannot activate proproliferative genes Rb-P Rb + E2F CyclinD.cdk4 CyclinD Rb: E2F P16 Ink4A MAPK pathway Ras
p53 is a transcriptional activator: One of the genes induced by p53 is p21, an inhibitor of the cdk4 kinase activity p27 Rb captures E2F, so that it cannot activate proproliferative genes CyclinE.cdk2 Rb-P Rb + E2F p53 CyclinD.cdk4 p21 CyclinD Rb: E2F p16 Ink4A MAPK pathway Ras
DNA damage leads to a block in cell cycle progression Replication of damaged DNA would fix mutations for all daughter cells
Possible biochemical function of the Rad24 group of checkpoint proteins. Rad24, together with the four small subunits of RFC, is a component of a pentameric complex. By analogy with RFC, this complex might recognise the transition between ssDNA and dsDNA. Such a structure is produced by many repair pathways but the Rad24 complex may only efficiently recognise it in the context of repair complexes (not shown here). Once the Rad24 complex is bound, it then functions to recruit the ‘PCNA-like’ Rad17/Mec3/Ddc1 complex to the DNA, followed by additional recruitment of checkpoint proteins involved in signal transduction (e.g. Mec1 and Rad53)
Organisation of the DNA-damage-dependent checkpoint pathways of budding yeast. Two distinct types of DNA damage, in the context of repair complexes, are represented in the schematic. Some of the components of the NER complex, specific for UV photoproducts, are indicated. The pointers indicate the incisions generated by the structure-specific endonucleases ERCC1/XPF and XPG. The Rad50/Mre11/Xrs2 complex is involved in DSB repair. RAD9 and RAD24 define upstream ‘sensor’ branches of the pathway and seem to respond, primarily outside of S phase, to multiple types of DNA damage. Currently, no other members of the RAD9 branch have been identified but the RAD24 branch can be ordered genetically as shown although strict linearity of function within this branch is unlikely. Both the RAD9 and RAD24 branches converge on MEC1, a member of the PI3 kinase family. Within S phase, one or more independent pathways for sensing DNA damage exist. The S-phase-specific sensing pathways are also thought to converge on MEC1. The MEC1, RAD53 and CHK1 genes encode transducers of the checkpoint signal. These transducers regulate effectors that control the biological consequences, as indicated, of activation of this pathway. Effectors for Rad53 include: Dun1, involved in G2/M arrest and DDR induction; Swi6, encodes a transcription factor targeted by the G1 checkpoint); Pol -primase, a target of the S-phase checkpoint; and Cdc5, a ‘polo-like’ kinase which inhibits Clb/Cdc28 kinase. Chk1 seems to primarily regulate Pds1, a regulator of sister chromatid cohesion. There may be other MEC1-dependent, RAD53-independent effector pathways. Whether induction of efficient repair requires DDR induction is unclear but there is probably a contribution from post-transcriptional mechanisms.
Mammalian cells: The protein p53 is sensing DNA damage P53 has a very high turn-over DNA damage: p53 becomes phosphorylated and stabilized
The Events in p53 Activation DNA damage (indicated by the break in the double line at the top) is recognized by a "sensor" molecule that identifies a specific type of lesion and possibly by the p53 protein, using its C-terminal domain. The sensor modifies p53 (by phosphorylation) when both molecules correctly determine that there is damage. A modified p53 is more stable (enhanced half-life), and a steric or allosteric change in p53 permits DNA binding to a specific DNA sequence regulating several downstream genes (p21, MDM2, GADD45, Bax, IGF-BP, and cyclin G). Two modes of signaling for cellular apoptosis are possible: one requiring transcription and one involving direct signaling with no transcription of downstream genes required.
Cell cycle control in mono- versus multicellular systems • Monocellular systems: • Unlimited proliferation • Control by size, nutrients and sex • Multicellular systems: • Proliferation is limited to specific regions and circumstances: Growth factors, cell:cell-interactions, • In mammals growth and proliferation are independently regulated
Influence of cellular and viral proteins in the cell cycle machinery
Growth factor stimulation through membrane receptors Extracellular ligand binding domain Transmembrane domain Tyrosine kinase domain EGF receptor FGF receptor
Growth factor stimulation through a membrane receptor Phosphorylation causes Inactivation of Rb EGF-receptor Rb-P Rb E2F CyclinD.cdk4 Rb: E2F CyclinD Tyrosine kinase Rb captures E2F: E2F cannot activate proproliferative genes MAPK pathway Ras EGF
p16 Cycl D:CDK4 p15 Smads p27 RB Cycl E:CDK4 - p21 E2Fs Changes in Gene Expression Cell Proliferation (Cell Cycle) Cell growth inhibitors that act through a membrane receptor Anti-Growth-Factors e.g. TGFß TGFß-R
Introduction Current view: • accumulation of multiple mutations within genes of a single cell • mutations confer a competitive advantage for cell growth and (de-) differentiation • mutations lead to initiation and progression of malignancies
Proto-oncogenes • control cell proliferation and differentiation • are expressed in all subcellular compartments (nucleus, cytoplasm, cell surface) • act as protein kinases, growth factors, growth factor receptors, or membrane associated signal transducers
Oncogenes • Mutations in proto-oncogenes alter the normal structure and/or expression pattern • Act in a dominant fashion • gain of function