Download
1 / 1
10 likes | 122 Views
Method. AG NPP [t ha -1 ]. BG NPP [t ha -1 ]. total NPP [t ha -1 ]. Method. Description. Calculation. Reference. 2002. 2003. 2002. 2003. 2002. 2003. 1. 0,85. 1,32. 5,22. 8,11. 6,07. 9,43. Max (AG,) live biomass BG NPP is estimated by root/shoot ratio. 1. Peak live Biomass.

E N D
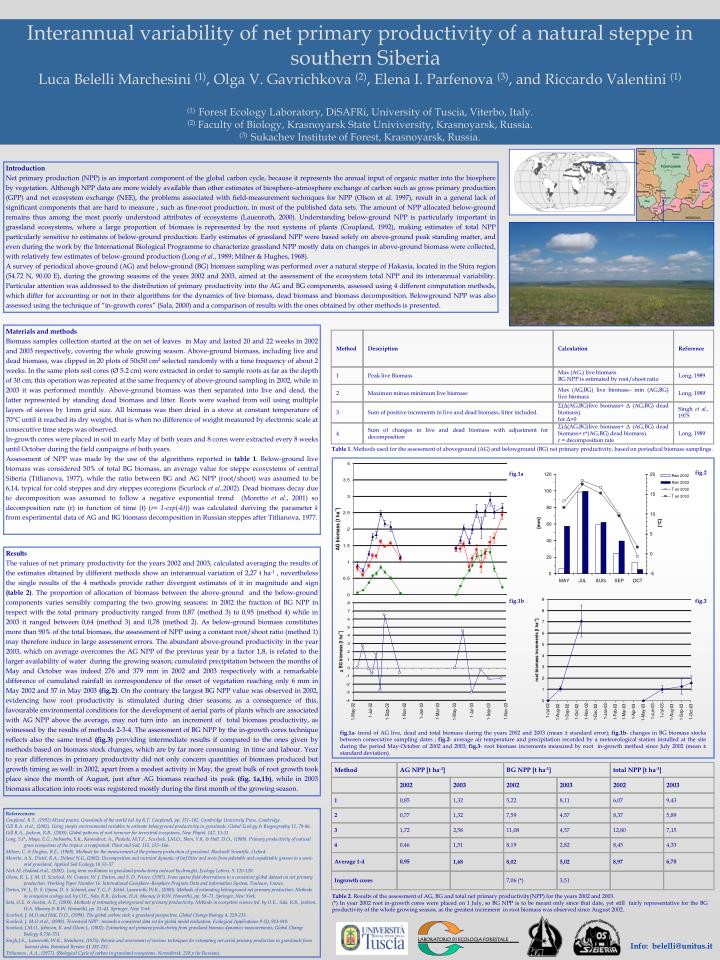
Method AG NPP [t ha-1] BG NPP [t ha-1] total NPP [t ha-1] Method Description Calculation Reference 2002 2003 2002 2003 2002 2003 1 0,85 1,32 5,22 8,11 6,07 9,43 Max (AG,) live biomass BG NPP is estimated by root/shoot ratio 1 Peak live Biomass Long, 1989 2 0,77 1,32 7,59 4,57 8,37 5,89 Max (AG,BG) live biomass– min (AG,BG) live biomass 2 Maximun minus minimum live biomass Long, 1989 3 1,72 2,58 11,08 4,57 12,80 7,15 Σ(Δ(AG,BG)live biomass+ Δ (AG,BG) dead biomass). for Δ>0 4 0,46 1,51 8,19 2,82 8,45 4,33 Singh et al., 1975 3 Sum of positive increments in live and dead biomass, litter included. Average 1-4 8,02 6,70 8,97 5,02 0,95 1,68 Σ(Δ(AG,BG)live biomass+ Δ (AG,BG) dead biomass+ r*(AG,BG) dead biomass). r = decomposition rate Sum of changes in live and dead biomass with adjustment for decomposition Long, 1989 4 Ingrowth cores 7,06 (*) 3,51 Introduction Net primary production (NPP) is an important component of the global carbon cycle, because it represents the annual input of organic matter into the biosphere by vegetation. Although NPP data are more widely available than other estimates of biosphere–atmosphere exchange of carbon such as gross primary production (GPP) and net ecosystem exchange (NEE), the problems associated with field-measurement techniques for NPP (Olson et al. 1997), result in a general lack of significant components that are hard to measure , such as fine-root production, in most of the published data sets.The amount of NPP allocated below-ground remains thus among the most poorly understood attributes of ecosystems (Lauenroth, 2000). Understanding below-ground NPP is particularly important in grassland ecosystems, where a large proportion of biomass is represented by the root systems of plants (Coupland, 1992), making estimates of total NPP particularly sensitive to estimates of below-ground production. Early estimates of grassland NPP were based solely on above-ground peak standing matter, and even during the work by the International Biological Programme to characterize grassland NPP mostly data on changes in above-ground biomass were collected, with relatively few estimates of below-ground production (Long et al., 1989; Milner & Hughes, 1968). A survey of periodical above-ground (AG) and below-ground (BG) biomass sampling was performed over a natural steppe of Hakasia, located in the Shira region (54.72 N, 90.00 E), during the growing seasons of the years 2002 and 2003, aimed at the assessment of the ecosystem total NPP and its interannual variability. Particular attention was addressed to the distribution of primary productivity into the AG and BG components, assessed using 4 different computation methods, which differ for accounting or not in their algorithms for the dynamics of live biomass, dead biomass and biomass decomposition. Belowground NPP was also assessed using the technique of “in-growth cores” (Sala, 2000) and a comparison of results with the ones obtained by other methods is presented. Materials and methods Biomass samples collection started at the on set of leaves in May and lasted 20 and 22 weeks in 2002 and 2003 respectively, covering the whole growing season. Above-ground biomass, including live and dead biomass, was clipped in 20 plots of 50x50 cm2 selected randomly with a time frequency of about 2 weeks. In the same plots soil cores (Ø 5.2 cm) were extracted in order to sample roots as far as the depth of 30 cm; this operation was repeated at the same frequency of above-ground sampling in 2002, while in 2003 it was performed monthly. Above-ground biomass was then separated into live and dead, the latter represented by standing dead biomass and litter. Roots were washed from soil using multiple layers of sieves by 1mm grid size. All biomass was then dried in a stove at constant temperature of 70°C until it reached its dry weight, that is when no difference of weight measured by electronic scale at consecutive time steps was observed. In-growth cores were placed in soil in early May of both years and 8 cores were extracted every 8 weeks until October during the field campaigns of both years. Assessment of NPP was made by the use of the algorithms reported in table 1. Below-ground live biomass was considered 50% of total BG biomass, an average value for steppe ecosystems of central Siberia (Titlianova, 1977), while the ratio between BG and AG NPP (root/shoot) was assumed to be 6,14, typical for cold steppes and dry steppes ecoregions (Scurlock et al.,2002). Dead biomass decay due to decomposition was assumed to follow a negative exponential trend (Moretto et al., 2001) so decomposition rate (r) in function of time (t) (r= 1-exp(-kt)) was calculated deriving the parameter k from experimental data of AG and BG biomass decomposition in Russian steppes after Titlianova, 1977. Table 1. Methods used for the assessment of aboveground (AG) and belowground (BG) net primary productivity, based on periodical biomass samplings. fig.2 fig.1a Results The values of net primary productivity for the years 2002 and 2003, calculated averaging the results of the estimates obtained by different methods show an interannual variation of 2,27 t ha-1, nevertheless the single results of the 4 methods provide rather divergent estimates of it in magnitude and sign (table 2). The proportion of allocation of biomass between the above-ground and the below-ground components varies sensibly comparing the two growing seasons: in 2002 the fraction of BG NPP in respect with the total primary productivity ranged from 0,87 (method 3) to 0,95 (method 4) while in 2003 it ranged between 0,64 (method 3) and 0,78 (method 2). As below-ground biomass constitutes more than 90% of the total biomass, the assessment of NPP using a constant root/shoot ratio (method 1) may therefore induce in large assessment errors. The abundant above-ground productivity in the year 2003, which on average overcomes the AG NPP of the previous year by a factor 1,8, is related to the larger availability of water during the growing season; cumulated precipitation between the months of May and October was indeed 276 and 379 mm in 2002 and 2003 respectively with a remarkable difference of cumulated rainfall in correspondence of the onset of vegetation reaching only 6 mm in May 2002 and 57 in May 2003 (fig.2). On the contrary the largest BG NPP value was observed in 2002, evidencing how root productivity is stimulated during drier seasons; as a consequence of this, favourable environmental conditions for the development of aerial parts of plants which are associated with AG NPP above the average, may not turn into an increment of total biomass productivity, as witnessed by the results of methods 2-3-4. The assessment of BG NPP by the in-growth cores technique reflects also the same trend (fig.3) providing intermediate results if compared to the ones given by methods based on biomass stock changes, which are by far more consuming in time and labour. Year to year differences in primary productivity did not only concern quantities of biomass produced but growth timing as well: in 2002, apart from a modest activity in May, the great bulk of root growth took place since the month of August, just after AG biomass reached its peak (fig. 1a,1b), while in 2003 biomass allocation into roots was registered mostly during the first month of the growing season. fig.1b fig.3 fig.1a- trend of AG live, dead and total biomass during the years 2002 and 2003 (mean ± standard error); fig.1b- changes in BG biomass stocks between consecutive sampling dates ; fig.2- average air temperature and precipitation recorded by a meteorological station installed at the site during the period May-October of 2002 and 2003; fig.3- root biomass increments measured by root in-growth method since July 2002 (mean ± standard deviation). Referecences: Coupland, R.T., (1992) Mixed prairie. Grasslands of the world (ed. by R.T. Coupland), pp. 151–182. Cambridge University Press, Cambridge. Gill R.A. et al., (2002). Using simple environmental variables to estimate belowground productivity in grasslands, Global Ecology & Biogeography 11, 79-86. Gill R.A., Jackson, R.B., (2000). Global patterns of root turnover for terrestrial ecosystems, New Phytol. 147, 13-31 Long, S.P., Moya, E.G., Imbamba, S.K., Kamnalrut, A., Piedade, M.T.F., Scurlock, J.M.O., Shen, Y.K. & Hall, D.O., (1989). Primary productivity of natural grass ecosystems of the tropics: a reappraisal. Plant and Soil, 115, 155–166. Milner, C. & Hughes, R.E., (1968). Methods for the measurement of the primary production of grassland. Blackwell Scientific, Oxford. Moretto, A.S., Distel, R.A., Didoné N.G.,(2002). Decomposition and nutrient dynamic of leaf litter and roots from palatable and unpalatable grasses in a semi- arid grassland, Applied Soil Ecology 18 31–37 Nick M. Haddad et al., (2002). Long term oscillation in grassland productivity induced by drought, Ecology Letters, 5: 110-120 Olson, R. J., J. M. O. Scurlock, W. Cramer, W. J. Parton, and S. D. Prince, (1997). From sparse field observations to a consistent global dataset on net primary production. Working Paper Number 16. International Geosphere–Biosphere Program Data and Information System, Toulouse, France. Parton, W. J., D. S. Ojima, D. S. Schimel, and T. G. F. Kittel. Lauenroth, W.K., (2000). Methods of estimating belowground net primary production. Methods in ecosystem ecology (ed. by O.E., Sala, R.B., Jackson, H.A. Mooney & R.W. Howarth), pp. 58–71. Springer, New York. Sala, O.E. & Austin, A.T., (2000). Methods of estimating aboveground net primary productivity. Methods in ecosystem science (ed. by O.E., Sala, R.B., Jackson, H.A. Mooney & R.W. Howarth), pp. 31–43. Springer, New York. Scurlock, J. M.O and Hall, D.O., (1998). The global carbon sink: a grassland perspective, Global Change Biology 4, 229-233. Scurlock, J. M.O et al., (1999). Terrestrial NPP : towards a consistent data set for global model evaluation, Ecological Applications 9 (3), 913-919. Scurlock, J.M.O., Johnson, K. and Olson J., (2002). Estimating net primary productivity from grassland biomass dynamics measurements, Global Change Biology 8,736-753. Singh,J.S.,, Lauenroth, W.K., Steinhorst, (1975). Review and assessment of various techniques for estimating net aerial primary production in grasslands from harvest data, Botanical Review 41 181-232. Titlianova , A.A., (1977). (Biological Cycle of carbon in grassland ecosystems. Novosibirsk, 218 p (in Russian). Table 2. Results of the assessment of AG, BG and total net primary productivity(NPP) for the years 2002 and 2003. (*) In year 2002 root in-growth cores were placed on 1 July, so BG NPP is to be meant only since that date, yet still fairly representative for the BG productivity of the whole growing season, as the greatest increment in root biomass was observed since August 2002. LABORATORIO DI ECOLOGIA FORESTALE Interannual variability of net primary productivity of a natural steppe in southern Siberia Luca Belelli Marchesini (1), Olga V. Gavrichkova (2), Elena I. Parfenova (3), and Riccardo Valentini (1) (1) Forest Ecology Laboratory, DiSAFRi, University of Tuscia, Viterbo, Italy. (2) Faculty of Biology, Krasnoyarsk State Univiversity, Krasnoyarsk, Russia. (3) Sukachev Institute of Forest, Krasnoyarsk, Russia. Info: belelli@unitus.it