Download
1 / 33
330 likes | 462 Views
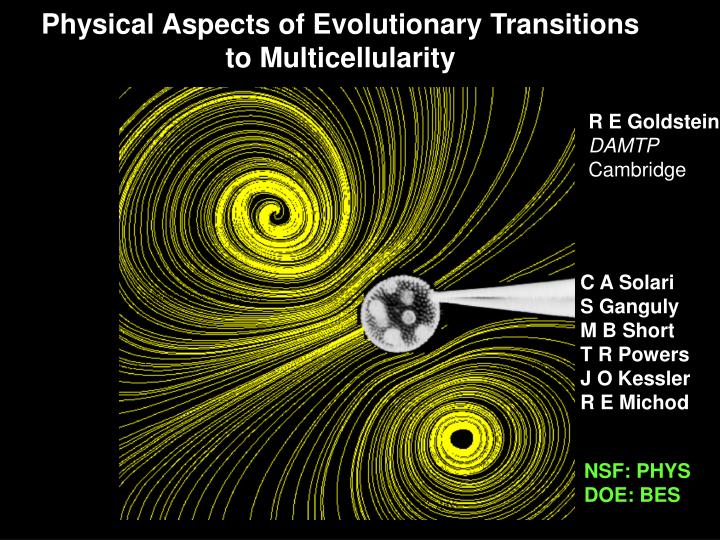
Physical Aspects of Evolutionary Transitions to Multicellularity. R E Goldstein DAMTP Cambridge. C A Solari S Ganguly M B Short T R Powers J O Kessler R E Michod. NSF: PHYS DOE: BES. Multicellularity. Uni- to multicellular organisms. Populations of individuals. Cellularization.

E N D
Physical Aspects of Evolutionary Transitions to Multicellularity R E Goldstein DAMTP Cambridge C A Solari S Ganguly M B Short T R Powers J O Kessler R E Michod NSF: PHYS DOE: BES
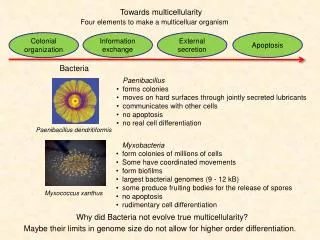
Multicellularity Uni- to multicellular organisms Populations of individuals Cellularization Chemotaxis & quorum sensing D. Mandoli (U. Washington) Park, et al. (2003) V. harveyi Acetabularia crenulata E. coli Signaling, adaptation, chemotaxis, (e.g., Berg & Purcell) Advantages of size, complexity, differentiation, transport (e.g., Bonner, Niklas)
(D.L. Kirk) Some Terminology Homocytic – 1,2, or 3-dimensional assembly of prokaryotic or eukaryotic cells that are structurally and functionally equivalent Heterocytic – differentiation of structure/function Colonial – physical association without cytoplasmic connections Multicellular – … with cytoplasmic connections Colonialism need not precede heterocytic lifestyle Lee, Cox, G (1996) D. discoideum
Volvox as a Model Organism Weismann (1892), Kirk (1998) • Extant collection, spanning from • unicellular to differentiated multicellular • Readily obtainable in nature, cultured • under a wide range of conditions • Studied from differing perspectives • (biochemical, developmental, genetic) • Broad ecological studies, with information • on distribution, environmental effects • Recent enough that genome may retain • traces of genetic changes in organization • Evidence of repeated genetic changes, • with hope that key ones are modest • Amenable to modern molecular-genetic • methods, such as DNA transformation van Leeuwenhoek (1700) Named by Linnaeus (1758) And, for theorists, it is the proverbial “spherical cow”!
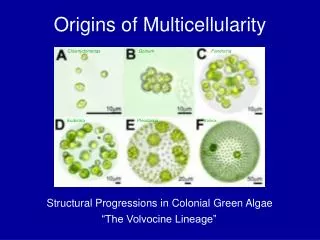
Chlamydomonas reinhardtii Eudorina elegans Gonium pectorale A Family Portrait Pleodorina californica Volvox carteri Volvox aureus somatic cells daughter colonies Germ-soma differentiation
Life Cycles of the Rich and Famous division Maturation of gonidia inversion hatching of juveniles cytodifferentiation and expansion
Germ-Soma Differentiation: regA gene In e.g. Chlamydomonas, the “ancestral” life cycle is: vegetative → reproductive → vegetative Palintomy: reproductive cells first grow and then divide by multiple fission. 8 colonies 8 cells d=3 divisions Grows 2d In e.g. Volvox, there is terminal differentiation, and after birth of daughter colonies, somatic cells undergo programmed cell death (apoptosis) “Somatic regenerator” mutants (R. Starr, 1970) led to discovery that there Is a single gene (regA) whose mutation gives rise Reg phenotype, in which somatic cells spontaneously revert to reproductive ones. In other words, the role of regA in wild-type cells is to suppress all aspects of reproductive cell development. It is off in gonidia, on in somatic cells.
Advection, Dissipation & Diffusion: Reynolds and Peclet Numbers Navier-Stokes equations: Passive scalar dynamics: Reynolds number: Peclet number: If U=10 mm/s, L=10 mm, Re ~ 10-4, Pe ~ 10-1 At the scale of an individual bacterium, dissipation dominates inertia, and advection dominates diffusion. The second relation breaks down with multicellularity…
The Diffusional Bottleneck Metabolic requirements scale with surface somatic cells: ~R2 Diffusion to an absorbing sphere Currents PO42- and O2 estimates yield bottleneck radius ~50-200 mm (~Pleodorina, start of germ-soma differentiation) Organism radius R
Source-Sink Hypothesis (Bell & Koufopanou, ’85,’93) Biological Considerations • The extracellular matrix is a source of nutrients, germ cells are sinks. • this enhances nutrient uptake rates over that possible with • isolated germs cells. • experimental demonstration – liberated germ cells grow more slowly • than those within ECM Flagellation Constraint Anchoring structures of flagella (“basal bodies”) serve as microtubule organizing centers (MTOCs) during cell division Hence, flagella beating stops during cell division, the time of high metabolic activity Other aspects of Flagella/Cilia/Undulipodia Left-Right symmetry breaking in embryonic development, mucus clearing, oocyte transport, kidney function, eyes, … (Ibanez-Tallon, et al., 2003) Highly-conserved nature of proteins from Chlamydomonas to humans (Pazour, et al., 2005)
Broken Colonies Deflagellated Colonies Flagellated Colonies Stirred, not Shaken Colchicine, a flagellar regeneration inhibitor (binds to tubulin, prevents microtubule polymerization) [Solari, Ganguly, Michod, Kessler, Goldstein, PNAS (2006)] Consistent with “Source-sink hypothesis” Bell & Koufopanou (‘85,’93) Inhibitor of flagella regeneration Deflagellation +Inhibitor Still medium Deflagellation +Inhibitor Bubbled medium Liberated +Inhibitor Bubbled Liberated germ cells
Stirring by Volvox carteri Pseudo-darkfield (4x objective, Ph4 ring) micropipette
Measuring Volvox Flows Sujoy Ganguly, U. Arizona & U. Cambridge Physics Today, July 2006 (Backscatter) Time-exposure of Volvox carteri near a surface
A Closer View Fluorescence
Even Closer (Flagellar Motions Visible) Fluorescence
Even Closer (Locally Chaotic Advection) Fluorescence
High-Speed Movie (125 fps) of Volvox Flagella Phase contrast +/- cytoplasmic connections between somatic cells: same flagellar coordination (Hiatt & Hand, 1972)
Fluid Velocities During Life Cycle Solari, et al. (2006) Daughter Division Hatch Pre-Hatch
Peclet Number During Life Cycle Division Pre-hatch Daughter Hatch This is “Life at High Peclet Numbers”
(Or, how to measure the average force per flagellum) Biological Version of Millikan Oil Drop Experiment Colonies are pH sensitive – they will throw off their flagella when the pH is transiently lowered - regrowth takes 90 minutes Regrowth can be in inhibited with colchicine (binds to tubulin)
Flagellar-Driven Flows and Scaling Laws Specified shear stress fat surface Larger colonies swim faster Measure <f> by deflagellation expt. Free-swimming colonies: Colonies held in place:
Velocity Profile (Experiment & Theory) Short, Solari, Ganguly, Powers, Kessler & Goldstein, PNAS (2006)
Metabolite Exchange Acrivos & Taylor (1962) – heat transport from a solid sphere: Magar, Goto & Pedley (2003) – prescribed tangential velocity in a model of “squirmers” Near surface: Boundary layer scale: In dimensionful terms, the boundary layer is Ra
Finite-Element Calculations Boundary Layer Scaling The Peclet number scales as: “Solute plumes” – like those of marine snow (Kiørboe & Jackson, 2001) (Roman Stocker, MIT – microfluidic studies of these plumes)
Phenotypic Plasticity I. Q: If colonies are deprived of nutrients, how do they adjust? A: By growing larger (!) Still medium Bubbling medium
Phenotypic Plasticity II. Q: If colonies are deprived of nutrients, how do they adjust? A: By growing longer flagella and beating them faster (!) Up velocity (still) Up velocity (dilution) Beating rate (dilution) Flow rate/10 (dilution) Flagella length (still) Flagella length (dilution)
Some Next Steps Further test of the scaling laws rotational frequency, swimming velocity Direct measurements of photosynthesis with/without flagella-driven stirring Flagella synchronization via hydrodynamic coupling a general problem involving molecular motors phototactic steering via a network of somatic cells