Download
1 / 50
520 likes | 695 Views
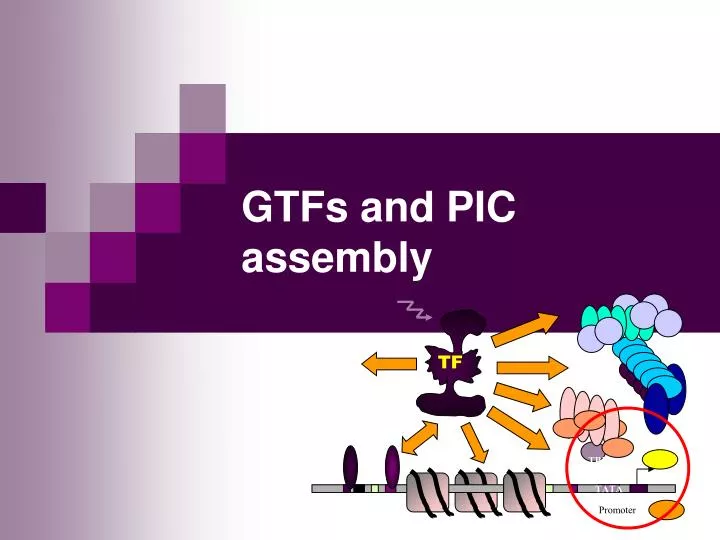
TF. TBP. TATA. Promoter. GTFs and PIC assembly. TBP. TFIIB. TFIIA. TFIIE. TFIIF. TFIIH. GTFs and PIC assembly. General transcription factors (GTFs) make RNAPII capable of selective initiation in vitro Highly conserved RNAPII+GTFs = ca. 30 polypeptides ≈ 2 MDa. +. =. PIC.

E N D
TF TBP TATA Promoter GTFs and PIC assembly
TBP TFIIB TFIIA TFIIE TFIIF TFIIH GTFs and PIC assembly • General transcription factors (GTFs) • make RNAPII capable of selective initiation in vitro • Highly conserved • RNAPII+GTFs = ca. 30 polypeptides • ≈ 2 MDa + = PIC Correct initiation of trx in vitro
Linear assembly of PIC -the preinitiation complex • Nucleation • TFIID+TATA form an “initial committed complex” • TAFs + INR may also initiate PIC-assembly • Common: a core sequence is recognized by a seq.spes.GTF • Link • initial complex recognized by TFIIB • With TFIIB bound, the complex becomes accessible to RNAPII • RNAPII recruitment • Assembly of RNAPII assisted by TFIIF • Minimal initiation complex formed • Maturationto complete trx competent PIC • Minimal initiation complex (DABF-pol) NOT trx.competent • Recruitment of TFIIH and TFIIE necessary • This step is unique for RNAPII
Alternatives to linear PIC-assembly Alternative Nucleation events Nucleation Link RNAPII recruitment Holoenzyme 2-step alternative Maturation
TBP [TFIID] function • Binds TATA - main sequence recognition event during PIC assembly • Binds a variety of different TATA-like sequences • A slow binding reaction • minor groove contact • binds as monomer • Affinity of TBP for TATA contributes to promoter strength • Binds also several other polypeptides • activators (Sp1, Tax1, E1A) • TAFs (dTAF110, dTAF40) • GTFs (TFIIB, TFIIA) • inhibitors • TBP = universal TF involved in all three pol syst. • TBP i SL1, TFIID, TFIIIB Other factors N DNA
TAFs TBP TBP versus TFIID • Subunit-structure • TFIID = TBP + multiple TAFs • mammalian TFIID: 750 kDa (II), 300 kDa (III) and 200 kDa (I) • TBP only a small core in the TFIID complex • human 38 kDa, yeast 27 kDa, Arabidopsis 22 kDa • TBP = N-term divergent domain + C-term. conserved domain • C-term domain 180aa symmetric • Carries all essential functions • N-term domain divergent • probably involved in regulating DNA binding TBP TFIID N
TBPs saddle-structure Convex surface protein Concave inside DNA Stigbøyler stirrups • 3D: saddle-structure • Twofold symmetry -form of a saddle. • Concave inside binds DNA in minor groove through a 10-stranded antiparallel -sheet • Convex surface binds other GTFs via 4 -helixes • loop (“stirrup”) on each side with Phe side-chains intercalating in DNA
.. but this way Not like this TBPs effect on DNA • DNA-structure is distorted upon TBP binding • DNA severely bended, unwinded and distorted • DNA shaped by TBP´s -sheet • The intercalating Phe-residues contributes to kink • Effect? • Upstream and downstream elements brought closer together • incompatible with nucleosome structure
A Two-Step Mechanism of TBP Binding to DNA • First step • Full-length TBPWT first binds to TATA box to form an unbent TBP-TATA box complex. • Second step • Then, this unbent complex slowly forms the bent TBP-TATA box complex. • TFIIB can directly recognize the unbent and/or bent TBP-TATA-complexes to form the bent TBP-TATA box complex. • Does this two-step mechanism apply within the context of TFIID?
TFIIB • Functions in PIC-assembly as adaptor - a molecular bridge that couples TBP-TATA with RNAPII • TFIIB recognizes the distorted TBP-TATA complex • contacts DNA on both sides of TBP-TATA • upstream via major groove (BRE) and downstream via minor groove • Provides directionality to the complex through assymmetric binding • TFIIB mediates RNAPII binding • interaction also with TFIIF • Function in initiation: “Measures” distance TATA - TSS TATA BRE TFIIB +1 TSS
TFIIB • TFIIB also contact point for activators • VP16, Steroid hormone receptorer, fushi tarazu, TAF40 • TFIIB-BRE: a repressive interaction? • The BRE was recently reported to repress basal transcription, with activator-mediated disruption of the BRE-TFIIB interaction as a proposed mechanism of gene activation. BRE TFIIB +1
TFIIB-structure • C-terminal core domain (cTFIIB) • C-term core with 2 imperfect direct repeats (2x 75aa) that binds TBP-TATA complex • each repeat = 5 -helices compact globular domain (cyclin A-like) • HTH motiv that binds BRE (not conserved in yeast and plants) • DNA-contact before and after TBP • N-terminal (nTFIIB) essential for RNAPII contact • cysteine-rich region that forms a “zinc-ribbon” + B-finger • mediate contact wtih RNAPII-TFIIF complex through a penetration mechanism C-term core TFIIB +1 N
TFIIBc structure TBP Two globular repeats contact DNA before and after TBP TSS TFIIB
TFIIB links TATA and RNAPIIand penetrates the active site TBP BC link TATA-pol TFIIB The N-terminal domain of TFIIB (Zn ribbon) binds the dock domain, where its B-finger plunges down into the RNAPII active center, loops back and remerges across the saddle. BN active site
TFIIB-B-finger penetrates RNAPII Boeger, H., Bushnell, D.A., Davis, R., Griesenbeck, J., Lorch, Y., Strattan, J.S., Westover, K.D. and Kornberg, R.D. (2005) Structural basis of eukaryotic gene transcription. FEBS Lett, 579, 899-903.
TFIIB-B-finger takes the place of RNAExpelled when trx starts B finger occupies the same location as the DNA–RNA hybrid. TFIIB may enhance the formation of an early transcribing complex before a length of 9 bp, required for optimal stability, is attained. As RNA grows, RNA and TFIIB must compete for space. If RNA wins, TFIIB is ejected and the pol is released from the promoter to complete trx of the gene. If TFIIB wins, initiation aborts and must be tried again. The B finger thus explains abortive initiation and promoter escape. TBP BC link TATA-pol TFIIB BN active site
Model for an RNAPII/IIF/IIB/TBP/DNA Minimal Transcription Complex
TFIIA • Controversial • not essensial in vitro with TBP and purified components • required with TFIID and less purified system • Function • counteracts repressors associated with TFIID (Dr1, topoI, MOT1) • Stabilizes the TBP-TFIIB complex • TFIIA needed for binding of TBP to nucleosomal TATA • TFIIA is able to enter the PIC assembly on all steps after TFIID binding • Required for activator-response
Structure of TFIIA • human/drosophila heterotrimer: 37 + 19 + 13 kDa () • Both and product of the same gene - the ab precursor is cleaved to a + b • yeast: heterodimer: 32 + 13 kDa • TOA1 32kDa (homologous to human and ) essensial • TOA2 13 kDa essensial • Antirepression requires • Activation requires • 3D two domains form an L-formed structure • TOA1 and TOA2 intertwined • Both C-terminals generate a compact -sheet ( -sandwich, -barrel) • Both N-terminals generate a “four-helix bundle” Yeast TOA1 Human a Human ß L C N
TFIIA structure C-terminal ß-barrel contacts DNA and TBP TFIIA N-terminal 4-helix bundle. Probably activator contact
TFIIA structure TBP C-terminal ß-barrel contacts DNA and TBP TFIIA N-terminal 4-helix bundle. Probably Activator contact
Yeast TFIIA + TBP + DNA TBP TFIIA
TFIIA - DNA-interaction • Interaction with DNA upstream TATA • C-terminal -barrel both TBP- and DNA-interaction • TBP-TFIIA: the edges of the two -structures interact extended -sheet • DNA-TFIIA: C-terminal -barrel contacts phosphates 3 bp upstream TATA • Explains why TFIIA stabilizes TBP-DNA complex • TFIIAs N-terminal -helix structure generates an interaction domain necessary for activator contact • Rational explanation of: • Antirepression requires which generate -barrel with TBP+DNA contact • Activation requires which also generate the N-terminal interaction domain • TFIIA and TFIIB bind on opposite sides of DNA without collision • TBPs convex surface still exposed for other interactions
TFIIA-TBP-TFIIB: place for all TBP TFIIA TFIIB
TFIIF (also called RAP = RNAPII-ass. faktor) • Structure: • Heterodimer in higher eukaryotes: RAP30 + RAP74 (Mw: 26 + 58 kDa) • S.cer.TFIIF heterotrimer: 105, 54, 30 kDa • Distinct feature: function in initiation and elongation • Initiation - helps in the recruitment of RNAPII • Stable association of RNAPII requires TFIIF • TFIIF-TFIIB associate in solution • TFIIF-RNAPII associate in solution • Initiation: a role in recruitment of TFIIE+TFIIH • Elongation: enhances catalytic velocity of RNAPII • More later TFIIB TFIIF
TFIIF = heterotetramer (RAP302 RAP742) RNAPII • RAP30: Two -related domains • RAP74: • Required for stimulation of elongation • RAP74 is strongly phosphorylated in vivo • Kinase? Possibly TAFII250 • TFIIF becomes more active when phosphorylated 30 30 DNA TFIIB 74 74 P P P P DNA RNAPII
TFIIF DNA-contacts • Complex pattern of protein-DNA contacts • Explained by wrapping of DNA around RNAPII-TFIIF? 74 74 30 30 ? TATA INR
3D of TFIIF • TFIIF (blue) is distributed across the surface of the polymerase. • The distribution of the second largest subunit of TFIIF was very similar to the sigmal subunit of bacterial RNA polymerase.
TFIIE TFIIEb • Structure • heterotetramer 22: 56 + 34 kDa • Contacts DNA in and just downstream of trx bubble • Function in trx.initiation • Recruitment of TFIIH to PIC • Regulates the activity of TFIIH • Role in NER (nucleotide excision repair) • Damage recognized by XPA • XPA binds TFIIE • TFIIE recruits TFIIH • Repairosome is formed 34 34 56 56 TFIIEa
TFIIH • The most complex of the GTFs - 9 subunits • Alle tilhørende gener essensielle • The only GTF with enzymatic activity: • Two Helicases (ATP-dep.) • [ATPase (DNA-dep.)] • CTD-kinase • Kinase substrat: • CTD - preferred substrate of Holo TFIIH • GTFs • TBP • TFIIEa • TFIIFa (RAP74) • Andre TFs • Oct, p53, RARa, ERa, pRb
core kinase TFIIH-structure Helicases utilise the energy of nucleotide hydrolysis to unwind nucleic acid duplexes. NER - nucleotide excision repair Surprising Link to DNA-repair • Multisubunit factor( human / yeast ) • 89 kDa XPB / SSL2 (p105) NER-function ATPase/3´-5´-helicase • NTP-site mutated lethal + trx.dead • XPB-helicase is necessary for trx.activity • Explains ATP requirement in initiation of trx • 80 kDa XPD/ RAD3 (p85) NER-function ATPase/5´-3´-helicase • NTP-site mutated not lethal + trx.OK + NER-defect • XPD-helicase not required for trx. activity • 62 kDa P62 / TFB1 (p75) UV-hypersens. • 50 kDa P52/ TFB2 (p55) • 44 kDa P44 / SSL1 (p50) (supr. of stem-loop) zinc finger motif • 34 kDa P34 / TFB4 (p37) zinc finger motif • 32 kDa MAT1 / TFB3 (p38) ring finger motif, cdk-assembly factor • 38 kDa cyclin H / CCL1 (p45+p47) cyclin-partner for CDK7/MO15 and Kin28 • 40 kDa CDK7, MO15 / KIN28 (p32) cyclin-dependent kinase • TFIIH dual function: in transcription initiat. and in NER
Core TFIIH Kinase (CAK) CAK Holo TFIIH = core TFIIH + CAK linked by XPD Bridge
TFIIH multiple functions • Function 1: promoter-melting assisted by helicases (2 steps, see below) • Model: 3´-5´-helicase + 5´-3´-helicase + ATP chain separation around TSS • ATP-depent step in initation (in addition to CTD phosphorylation) • Function 2: CTD-kinase, role in promoter clearance • Modell: CTD-phosphorylation after chain separation and initiation PIC disrupted elongation complex leaves the promoter • Function 3: role in elongation • Model: TFIIH-kinase+ ATP maintains hyperphosphorylated pol.II (counteracting the CTD phosphatase) • Function 4: role in DNA-repair (NER) • 5 of 9 subunits of TFIIH with a double function in trx.+repair • actively trx.genes are preferentially repaired • TFIIH can complement NER-deficient extract
Assists in formation of open complex and promoter escape 1. ATP-dependent promoter melting - chain separation - open trx. complex 2. ATP-dependent structural transition into an escape- competent conformation TFIIH helicase TFIIH helicase
TFIIH: also linked to the cell cycle? • The TFIIH kinase = CAK = cdk7 + cyclin H + MAT-1 • CAK = CDK activating kinase • CAK activates other cdk´s through Thr-phosphorylation • MAT-1 (a ring-finger protein) makes CAK constitutively active (Thr-indep.) • An open question : • Is CTD-phosphorylation regulated by the cell cycle? • Different answers : • No - probably not! • Argument : only 20% of all CAK in the cell is TFIIH-associated • S.cer. Has separate CAKs for TFIIH and cell cycle • Activity and level of CDK7, cyclin H and Mat1 do not change during cell cycle • Yes - May wll be! • TFIIH inhibited during mitosis concomitant with inhibition of CDK7 (CDC2-induced) • Cell cycle inhibitor INK4 inhibits CTD phosphorylation by CDK7 • CDK8 can negatively regulate CDK7
Model Repair- proteins DNA repair Transcription CAK CAK Core TFIIH Core TFIIH CAK Cell cycle
TBP TBP TFIIB RNAPII TBP TFIIB RNAPII TFIIF TFIIE PIC assembly - a gradual wrapping process?
Multiplicity of GTFs? Are a single set of GTFs universally used? …equally at all promoters?
Several GTF complexes possible • Several GTFs encoded by single copy genes • TFIIB, E, F, and H • Also true for RNAPII • However, multiple genes exist for specific GTFs • Multiple TFIIA related • Multiple TFIID related • Gene-selective developmental roles? • Consequence: several possible complexes possible • By replacing ”normal” versions with specific ones • By generating variant combinations of GTF-containing complexes
Variant TBPs:TRFs = TBP related factors ≥2 TBP like proteins in multicellular organisms TBP top view Drosophila TRF1 TBP TRF2 TLP TLF TRF TRP ≈ TBP bottom view • TRF1 - major part of TFIIIB, a RNAPIII factor • TRF1 binds pref TC-box (TTTTCT) in the core promoter of the Drosophila tudor gene, a direct target TBP specific TRF2 specific
A diversity of complexes • Many TBP complexes • Alternative TAF-containing complexes • Variant TFIIAs
A diversity of core promoters may assemble gene-specific complexes • TATA core promoters require TBP, but not necessarily TAFs • Inr ± DPE core promoters require TAFs and hence indirectly TBP associated • TLF-dependent core promoters do not require TBP
Enormous diversity Diversity of core promoters • GTF machinery shows some diversity • Activators and repressors (Tfs) show enormous diversity • Not thousands to one, but thousands to several Some diversity